毛竹邻近林分结构与立地对其自然扩张的影响
2019-04-23王秀云宋绪忠庞春梅赵明水
王秀云,宋绪忠,庞春梅,杨 华,赵明水,徐 良
毛竹邻近林分结构与立地对其自然扩张的影响
王秀云1,宋绪忠1,庞春梅2,杨 华1,赵明水2,徐 良2
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江天目山国家级自然保护区管理局,浙江 杭州 311311)
为探明毛竹林自然扩张的影响因素及程度,2005年在浙江省天目山国家级自然保护区选定9个毛竹纯林小班,在其四周共设置36个样地,测定分析样地林分的平均胸径、平均树高、郁闭度、林龄、林分密度、树种组成、坡度、坡位、土层厚度及林分起源10个因子,2015年复测,分析10 年间毛竹的自然扩张距离和10因子的相关关系。主成分分析结果表明,反映林分密度、平均树高、平均胸径、郁闭度、树种组成的第一个主成分因子方差贡献率最高,达46.99%,对毛竹林自然扩张的影响最大;表征坡度、坡位及土层厚度的第二个主成分因子,方差贡献率为20.26%,对毛竹自然扩张的综合影响较第一个主成分因子小;第三个主成分因子是林分年龄和起源,对毛竹林自然扩张的影响最小。分析结果显示,邻近森林群落林分结构与立地条件决定了毛竹扩张能力,邻近群落结构简单,物种多样性低,其抗干扰能力就低,毛竹容易入侵。
毛竹;自然扩张;林分结构;立地条件
毛竹属禾本科Gramineae竹亚科Bambusoideae刚竹属,是一种高大散生状乔木型植物,主要分布在我国亚热带地区[1]。它依靠竹鞭强大的繁殖和空间拓展能力,不断向邻近森林群落扩张、发笋成竹,形成竹阔混交林并不断纯林化[2]。毛竹林不断无序扩张,给邻近的森林群落构成了巨大威胁,因其改变群落结构和组成、降低动植物生物多样性[3]、抑制群落自然更新、减弱生态系统功能、破坏森林土壤理化性质与微生物组成[4]等,已引起各级政府、生态学与林学研究者的广泛关注。
毛竹能向邻近森林群落扩张,既受自身生物属性影响,也与邻近森林生物与非生物因素(可侵入性)有关[5]。然而对毛竹扩张的研究多集中在其产生的影响方面,对毛竹扩张的影响因素研究鲜有报道。林分结构既是评价森林健康状况、稳定性的重要指标,更是衡量森林抗干扰和自我修复能力的关键因子,同时,森林生长状况受立地因子影响明显。因此本研究在浙江省天目山自然保护区内,选择少受人为干扰的毛竹林为对象,深入分析毛竹邻近森林群落林分结构与立地条件对毛竹扩张的影响,明确影响毛竹自然扩张的主要因素及影响程度,以期能揭示毛竹自然扩张规律,同时对提前防控毛竹扩张产生的生态影响及恢复和管理受侵森林群落或生态系统均具有重要意义。
1 研究区概况
研究区设在浙江省天目山自然保护区,191°24' 11"191°27' 11" E,30°18' 30"30°21' 37" N。年平均气温8.915.8°C,最冷月气温2.63.4°C,极端最低气温-20.2°C,最热月气温19.928.1°C,≥10°C年积温2 5005 100°C,无霜期209235 d;年降水量1 3901 870 mm,年太阳总辐射3 7704 460 MJ·m-2。属北亚热带气候,季风强盛,四季分明,气候温和,雨量充沛,光照适宜。地质古老,地形复杂,土壤肥沃,土壤基本属于亚热带红黄壤类型,适宜毛竹的生长发育。其森林植被大体划分为5个类型:常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、针阔混交林、黄山松纯林。天目山毛竹种群所处群落为竹阔混交林和阔叶林,层次明显,乔木层主要有青冈,苦槠,木荷,柳杉var. sinensis,杉木等;灌木层有毛柄连蕊茶,山矾,马银花和山胡椒等;草本植物荨麻,箬竹,一枝黄花,萱草等植物。天目山自然保护区自设立保护区后,禁止人为活动干扰,原本种植于山下的毛竹林迅速扩张,面积逐年增加。根据遥感影像数据,1984年、1994年和2015年,天目山毛竹林面积依次为146.16 hm2,224.82 hm2和381.78 hm2,30年间毛竹林面积增加了235.62 hm2,以平均7.6 hm2·a-1的速度迅速向山上蔓延,严重蚕食了周围的原始植被。
2 研究方法
2.1 样地设置与调查
2005年9月,在研究区选取9块典型未经人为干扰的毛竹纯林小班(表1),并在毛竹纯林小班外缘的上、下、左、右各设置1块样地,样地规格为60 m×60 m,共计36个样地,包括黄山松样地(4个)、松阔混交林(3个)、柳杉(11个)、杉阔混交林(10个)和阔叶林(8个),其中阔叶林(苦槠、青冈、木荷等)样地林分属天然林外,其余均为上世纪80年代初营造的人工林(表2)。然后将每个样地划分为5 m×5 m的样方,在每个样方中调查并记录胸径≥5 cm乔木的树种名、胸径、树高、相对位置坐标(,)及样地林分的郁闭度、土层厚度、坡度、坡位等信息,记录林下灌木层与草本层的植物种、平均高度、盖度及株数(丛数)。2015年9月同期复查样地,并记录样地内增加的毛竹株数、胸径、株高、年龄,并根据新增毛竹位置坐标计算距毛竹纯林小班边界的长度,以此确定为毛竹扩张距离;另外,有的毛竹已经穿过样地扩张到样地外,根据毛竹鞭根延伸方向,测量其距离毛竹纯林小班的长度,以此为扩张距离。
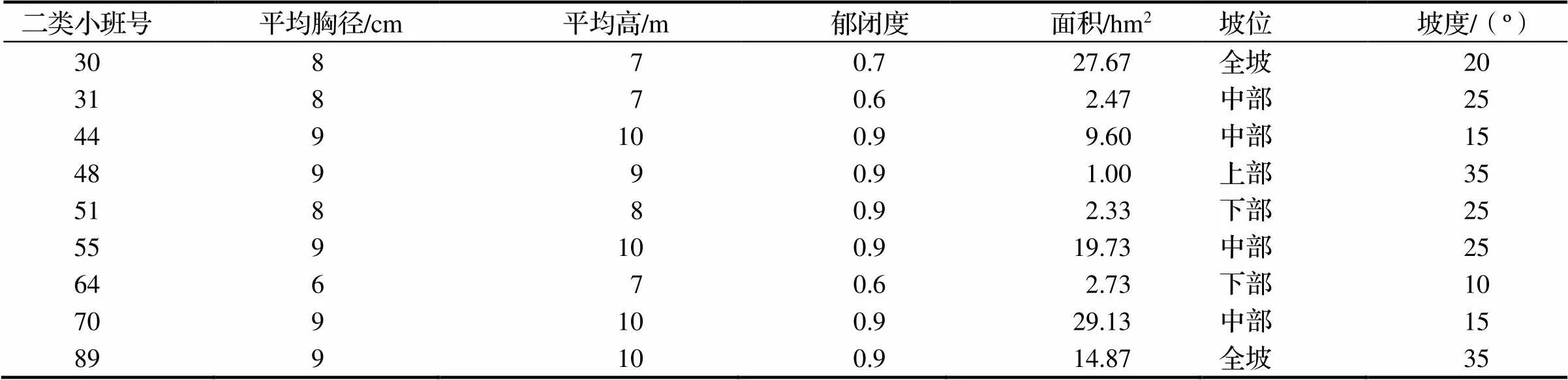
表1 毛竹纯林小班基本信息

表2 36个样地基本信息
2 主成分分析
2.1 影响因子调查
根据样地调查结果,选取林分结构指标平均胸径、平均树高、林分密度、郁闭度、龄组、林分起源、树种组成,及立地因子坡位、坡度、土层厚度,共10个因子作为毛竹扩张距离的影响因子,分析林分结构、立地因子对扩张距离的影响程度。
研究数据中,如林分起源、龄组、树种组成、坡位、土层厚度等因子是非数值型数据,需要将其转化为数值型数据,进行量化处理(表3)。

表3 数据量化方法
2.2 主成分提取
运用SPSS20.0的主成分分析方法[6-7],通过正交变换,在尽量不损失信息量的前提下,减少参评指标的原始变量个数,构建新的线性组合变量方程,同时可降低指标间的相关性,让参评因子间相互独立或不相关。数据分析时,特征根累积百分率反映所提取主成分对总方差的贡献率,特征向量代表各指标因子对主成分的贡献大小,可据此确定权重的大小。主成分提取时,提取样本方差累积贡献率一般情况下应大于80.00%,同时各主成分对应的特征值大于1。
采用SPSS软件中的主成分分析法研究毛竹入侵距离与林分结构、立地因子之间的耦合关系。通过降维,从众多的因素中找出主要的影响因素,用较少的变量来反映毛竹扩张能力。
2.3 主分量模型的建立
选择线性模型对主分量与林分结构、立地因子间的关系进行阐述,模型表达式如
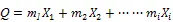
式中,为变量个数,即=1,2,3……;i为变量;m为变量系数。
各影响因子与主分量的关系可以通过其方差贡献率来解释,所用模型如下式表示:
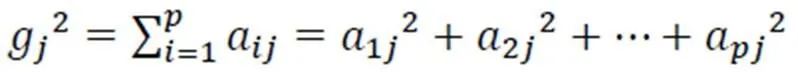
Q的贡献率公式如下:
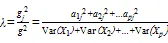
式中,Var()为原始变量的方差;a2为公共因子Q对于的分量X=(=1,2,……)的方差;为公共因子数;g2为公因子Q对原始变量的方差贡献;2为所有公因子对原始变量的方差贡献。
2.4 主成分综合得分
对于某一样地而言,各主成分因子得分和因子权重(主成份特征值)乘积之和便是主成分因子综合得分(Q),即样地的结构与立地综合指数,具体表达如下:
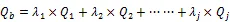
式中,Q为主成分因子,是主成份的特征值。
3 结果与分析
3.1 样地调查结果
36个样地调查发现,柳杉样地林分密度最小,平均450株·hm-2,黄山松最大,平均达2 190株·hm-2,其它样地密度1 1601 190株·hm-2。柳杉样地平均胸径44 cm,黄山松样地平均胸径最低,仅11 cm,其它样地林分平均胸径20 cm。样地林分下层分布有山矾、油茶、雀梅藤、山胡椒等,平均高度为1.5 m,盖度为80%;草本层主要有箬竹、荨麻、狗脊等,平均高度为0.5 m。
统计调查数据发现,10 年间毛竹向36个样地扩张距离的最小值是10 m,最大值达210 m,即扩张最快的毛竹已穿过样地,延伸到样地外,平均扩张距离为64.71 m。
3.2 主成分提取与表达
将平均胸径、平均树高、林分密度、郁闭度、龄组、林分起源、树种组成和立地因子坡位、坡度、土层厚度10个因子,利用SPSS 20.0统计分析软件,进行主成分分析。表4表明,前3个主成分的样本特征值均>1,且累积贡献率=81.43%>80%,符合主成分提取原则。
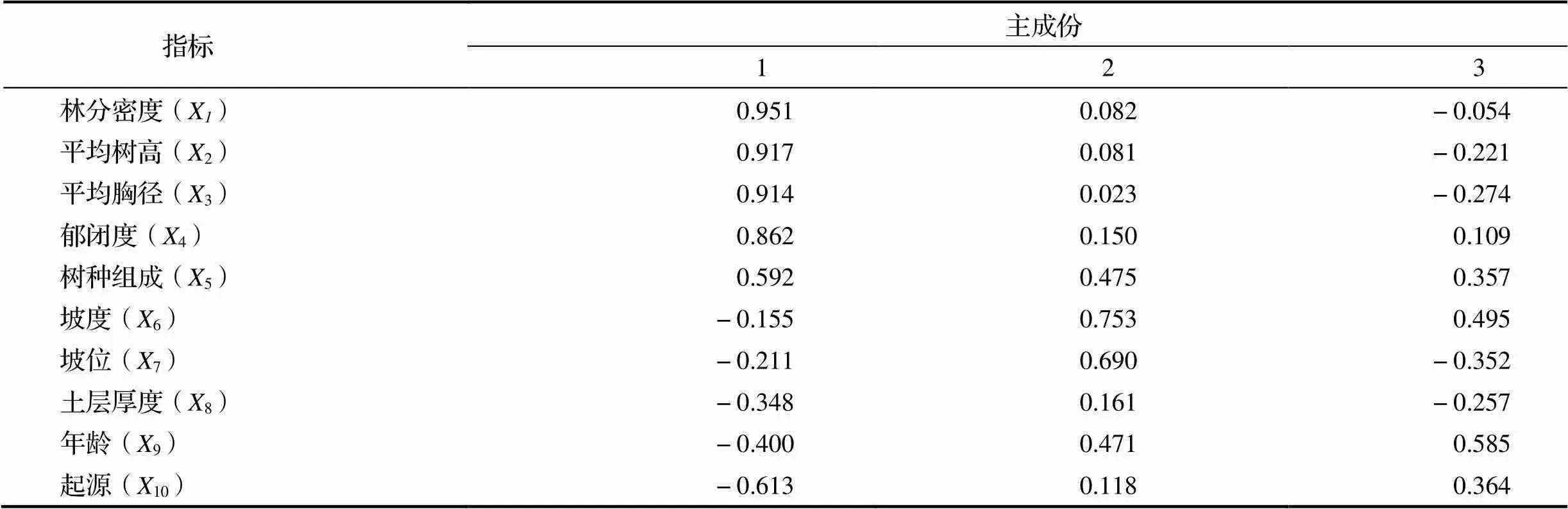
表5 因子载荷矩阵表
由表5可知,所提取出的3个主分量基本解释了10个因子的变异信息。其中第一主成分因子主要是林分密度、平均树高、平均胸径、郁闭度、树种组成的信息表达,其载荷量依次为0.951,0.917,0.914,0.862和0.592,表明第一主成分因子主要表达了林分结构的信息。第二主成分因子主要是坡度、坡位及土层厚度这3个指标,载荷量分别为0.753,0.690和0.161,表明第二主成分因子主要表达了立地条件的信息。第三主成分因子主要是年龄、起源,其载荷量是0.585和0.364,载荷量最小,贡献率最低。
3.3 主分量模型
根据主分量1,2及3的得分系数,可以列出由10个因子组成的主分量模型式(5)至式(7):

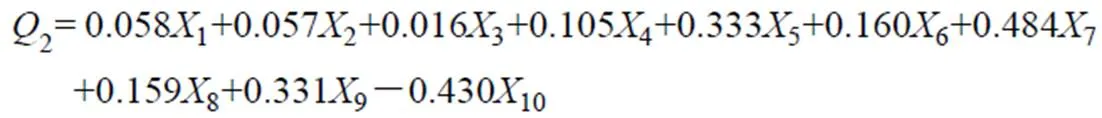
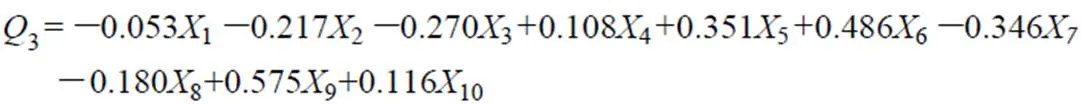
3.4 综合影响分析
主分量1,2,3与样地综合指数b与毛竹扩张距离()的关系散点图(图1)可发现,表征不同因子组合的主分量对毛竹林扩张距离的影响程度不同。表征林分结构因子的主分量1与毛竹扩张距离呈显著的对数关系,其相关系数达到0.903 4(<0.05)。毛竹邻近林分郁闭度、林分密度、平均树高、平均胸径越大,树种组成越复杂,其F1值越大,毛竹林向内扩张的距离越小。
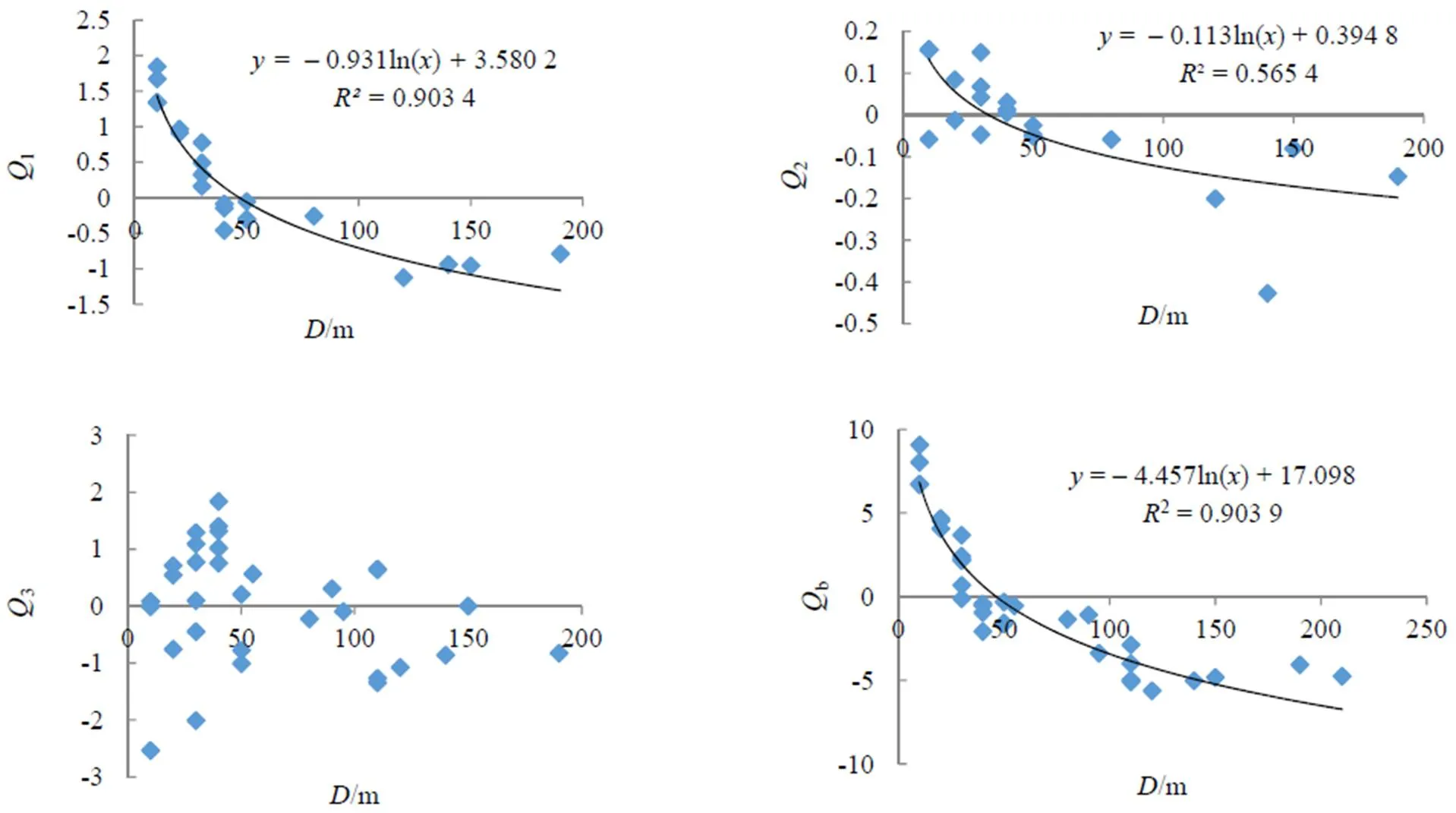
图1 毛竹林扩张距离与主分量Q1, Q2, Q3及综合指数Qb的相关关系
Figure 1 Correlation between principal components andcomposite index and spreading distance offorest
表征立地因子的主分量2与毛竹扩张距离的相关关系较显著,立地因子中影响毛竹扩张距离的是坡度,毛竹邻近林分所处立地越平坦,其2越小,毛竹扩张越容易,扩张距离越大。样地数据表明(图2),毛竹邻近林分坡度与扩张距离关系呈显著的相关关系,10º,15º,20º,25º,35º不同梯度坡度,毛竹扩张距离呈规律性下降。而表征林龄和起源的主分量3与毛竹扩张距离的相关性不显著。
样地综合影响评价指数b与扩张距离关系可用对数关系表达,其相关系数达到0.903 9(<0.05)。样地综合影响评价指数是样地林分结构与立地因子的信息表达,指数b越高表征该样地林分结构与立地条件越不利于毛竹林自然扩张,抗干扰能力越强;反之,指数越小,该样地林分结构与立地条件越利于毛竹林自然扩张。
4 结论与讨论
4.1 研究方法的讨论
主成分分析法用于研究毛竹林自然扩张的主要影响因素是可行的。从林分结构指标(包括平均胸径、平均树高、林分密度、郁闭度、龄组、林分起源、树种组成)及立地因子(包括坡位、坡度、土层厚度)共10个因子中寻找毛竹林自然扩张的3个主分量,既实现了数据简化的效果,又可更深入地理解各因子之间的相互关系,同时可揭示各主分量对毛竹自然扩张的影响程度。
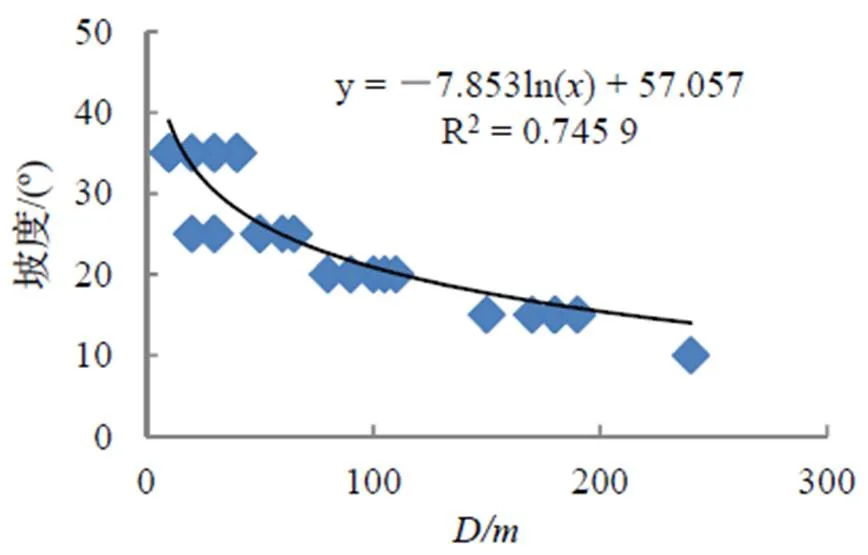
图2 毛竹林扩张距离D与坡度的相关关系
Figure 2 Correlation between slope and spread distance offorest
4.2 毛竹扩张的影响因素分析
影响毛竹林自然扩张的因素是多方面的,由多个影响因素相互联系、共同作用的结果。从主成分因子反映的信息来看,林分结构指标与立地因子均会产生影响,但其影响程度不同。林分密度、平均树高、平均胸径、林分郁闭度、树种组成、坡度对毛竹林自然扩张的影响大,而林分年龄、起源、坡位、土层厚度对其影响较小。
毛竹能否扩张,除与其自身的生长发育生物属性有关外,很大程度上依赖于邻近森林群落的抵抗力和稳定性,即群落可入侵性。邻近群落结构简单、物种多样性低,毛竹就容易入侵,正如本研究中,9块毛竹纯林小班位于自然保护区内,10年间未受到人为干预及经营,完全自然地向邻近林分扩张,如果邻近林分是郁闭度、林分密度较高的阔叶混交林,毛竹扩张的距离较小,而且有研究报道毛竹向灌木林、人工林和落叶阔叶林的平均扩张速率分别为3.0,2.5和1.8 m·a-1[8];同时毛竹扩张还受光热、土质、水湿条件、地形及海拔因素等的影响,如有研究报道毛竹扩张与群落所在坡度、坡向等有一定的相关性[9],而本研究的结果也证实了该论断。
4.3 毛竹扩张的潜力及产生的后果及对策讨论
毛竹扩张符合“内禀优势、资源机遇与干扰促进”的生物入侵机制[10-11],即毛竹依靠生长迅速、繁殖力强、表型多变、协同作用等自身生物优势,通过遮阴、机械损伤、抑制凋落物、竞争养分与化感作用等竞争方式,抑制其他植物生长并不断向邻近森林群落扩散蔓延,不同程度地影响了群落的结构组成、生物多样性、土壤理化性质与微生物组成、生态系统过程与功能等。据研究报道,天目山国家级自然保护区内毛竹林扩张迅速,1985-2003年毛竹林总面积增加了80.53 hm2,增加了3.4倍,平均增长4.47 hm2·a-1[12]。毛竹林的扩张改变了林地土壤有机碳的含量[13]、物种多样性[14-15]、土壤微生物组成[16]等生态问题,因此,有效防控毛竹无序扩张是一个非常有挑战性的问题。蔡亮等[1]提出了挖沟、灌水的设想,但在劳动力成本日益高涨的情形下,这个方法难以推行。应该因势利导,根据毛竹林经营目的,加强毛竹合理利用与精细化管理,通过增加采笋量抑制新增毛竹的进一步扩散;对壮龄期毛竹的扩张,适当疏伐毛竹,补植地带性阔叶树种或速生性树种;调整毛竹邻近森林群落结构,合理增加林分郁闭度和林分密度,构建稳定的针阔混交或阔叶混交复层林群落结构,加强其抗干扰能力,以抵御毛竹林自然扩张及其带来的影响。
[1] 蔡亮,张瑞霖,李春福,等. 基于竹鞭状态分析的抑制毛竹林扩散的方法[J]. 东北林业大学学报,2003,31(5):68-70.
[2] 潘璐,牟溥,白尚斌,等. 毛竹林扩张对周边森林群落菌根系统的影响[J]. 植物生态学报,2015,39(4):371-382.
[3] 白尚斌,周国模,王懿祥,等. 森林群落植物多样性对毛竹入侵的响应及动态变化[J]. 生物多样性,2013,21:288-295.
[4] 宋庆妮,杨清培,刘骏,等. 毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响[J]. 应用生态学报,2013,24:338-344.
[5] 杨清培,杨光耀,宋庆妮,等. 竹子扩张生态学研究:过程、后效与机制[J]. 植物生态学报,2015,39(1):110-124.
[6] 吴星. 统计原理中主成分分析法的应用[J]. 科研,2015,28:27.
[7] 高惠璇. 应用多元统计分析[M]. 北京:北京大学出版社,2005.
[8]Okutomi K,Shinoda S,Fukuda H. Causal analysis of the invasion of broad-leaved forest by bamboo in Japan[J]. J Veget Sci,1996,7:723-728.
[9] Suzuki S,Nakagoshi N. Expansion of bamboo forests caused by reduced bamboo-shoot harvest under different natural and artificial conditions[J].Ecol Res,2008,23:641-647.
[10] Blumenthal D M. Interactions between resource availability and enemy release in plant invasion[J]. Ecol let,2006,9:887-895.
[11] Catford J A,Jansson R,Nilsson C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework[J]. Divers Distr,2009,15:22-40.
[12] 丁丽霞,王祖良,周国模,等. 天目山国家级自然保护区毛竹林扩张遥感监测[J]. 浙江林学院学报,2006,3(3):297-300.
[13] 吴家森,姜培坤,王祖良. 天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响[J]. 江西农业大学学报,2008,30(4):689-692.
[14]林倩倩,王彬,马元丹,等. 天目山国家级自然保护区毛竹林扩张对生物多样性的影响[J]. 东北林业大学学报,2014,42(9):43-47,71.
[15] 杨淑贞,杜晴洲,陈建新,等. 天目山毛竹林蔓延对鸟类多样性的影响研究[J]. 浙江林业科技,2008,28(4):43-46.
[16] 王奇赞,徐秋芳,姜培坤,等. 天目山毛竹入侵阔叶林后土壤细菌群落16S rDNA V3区片段PCR的DGGE分析[J]. 土壤学报,2009,46(4):662-669.
Effects of Structure and Site Condition of Neighbor Forests on Natural Expansion of
WANG Xiu-yun1, SONG Xu-zhong1, PANG Chun-mei2, YANG Hua1, ZHAO Ming-shui2, XU Liang2
(1. Zhejiang Academy of Forestry, Hangzhou 310023, China; 2. Management Office of National Nature Reserve of Mount Tianmu of Zhejiang,Hangzhou, 311311,China)
In 2005, in Mount Tianmu National Nature Reserve of Zhejiang, 9 subcompartments were selected inedulis forest, and 36 sample plots were established around subcompartments for determination and analysis on mean DBH, tree height, crown density, forest age, density, species composition, slope, slope site, soil thickness and forest origin. In 2015, determinations were repeated. Principal component analysis on data of the 2 determination showed that the first principal component, including forest density, mean DBH, average height, crown density and species composition had contribution rate of 46.99%. The second principal component consisted of slope, slope site and soil thickness, had 20.26% contribution rate, indicating that site condition had lower effect than the first one on natural expansion of. The third principal component was about the age and origin of forest, had the lowest effect. The analysis resulted that expansion ofwas caused by simple structure and lower species diversity of neighboring forest.
; natural expansion; stand structure; site condition
S795.5
A
1001-3776(2019)06-0019-06
10.3969/j.issn.1001-3776.2019.05.004
2019-07-20;
2019-10-17
浙江省省院合作林业科技项目(2017SY09;2016SY15;2017SY12)
王秀云,副研究员,从事森林经理与森林生态研究;E -mail: lyhwxy@tom.com。
宋绪忠,研究员,从事森林生态与土壤修复研究;E -mail: popsong@163.com
