东北玉米根系生物量模型的构建*
2019-04-13吕国红谢艳兵温日红王笑影贾庆宇
吕国红, 谢艳兵, 温日红, 王笑影, 贾庆宇
东北玉米根系生物量模型的构建*
吕国红, 谢艳兵, 温日红, 王笑影**, 贾庆宇
(中国气象局沈阳大气环境研究所 沈阳 110166)
开展根系生物量的观测和研究, 建立通用性的根系生物量模型对于开展生态系统生物量的监测和评估具有重要意义。为得到根系生物量的实时信息, 2016年9月末利用挖土法和根系扫描系统, 获取玉米根系的生物量及生态指标, 分析了玉米根系生物量的垂直分布特征并建立了根系生物量与根系生态指标之间的模拟方程。结果表明: 玉米根系生物量主要集中于0~30 cm, 占玉米根系垂直分布量的94.44%。利用普通最小二乘法建立根系生物量模型均存在异方差问题, 增加根长作为自变量建立的根系生物量模型显著提高了模拟精度, 决定系数(2)达0.91以上。采用对数转换消除方程的异方差及比较不同的模拟方程后发现, 玉米根系生物量与根径和根长的组合变量(2)建立的指数函数是模拟玉米根系生物量的最优方程, 决定系数(2)最高, 为0.90, 平均绝对误差(MAE)、估计值的标准误差(SEE)、平均预估误差(MPE)均最小, 满足了模拟方程的精度要求。对该方程进一步验证发现, 模拟值和实测值之间的相关系数为0.92, 说明此模型能较好地模拟根系生物量。利用根系生物量模型结合微根管法, 可解决根系生物量实时观测难的问题。
玉米; 根系生物量; 根长; 根径
根系作为支撑、固定植物体的重要器官, 是植物吸收水分和养分的重要有机组成部分[1]。根系生物量是整个群落生物量的重要组成部分, 是评价植物生长能力的重要指标, 影响着群落的构建、结构和功能[2-4]。根系生物量占总生物量10%~20%, 足见其在物质循环和能量循环的重要性[5]。因此, 开展根系生物量的观测和研究, 建立通用性的根系生物量模型对于开展生态系统生物量的监测和评估具有重要意义。
植物根系大部分深埋于地下, 取样相对困难, 难以直接观测和测定, 导致研究根系生物量的深度和广度均落后于地上生物量[6], 因此, 建立根系生物量模型是估算根系生物量的有效方法。与地上生物量模型相比, 根系生物量模型数量相对较少, 如Zianis等[7]研究欧洲树种的607个生物量方程中, 只有11.5%的方程用于模拟地下生物量。根系生物量模型的建立多集中于森林生态系统, 如崔喜红等[8]利用探地雷达技术, 基于树木的根径估算根生物量; 曾伟生等[9]利用根茎比模型和地上生物量模型联合估计东北落叶松(spp.)和南方马尾松()的地下生物量; 肖义发等[10]利用树高、地径、胸径等建立了思茅松(var.)地下根系的主根、侧根和根系总生物量的模拟方程。由上可知, 国外也多是利用胸径、根茎比、树高等模拟森林地下生物量[11-13]。森林生态系统由于根系的难以获取, 多是利用地上生态指标模拟地下生物量, 较难实现根系生物量与根系生态指标直接建立模拟方程。农田生态系统较森林生态系统, 根系相对易获取, 建立根系生物量与根系生态指标的模拟方程相对较容易, 如刘永霞等[14]利用塑料桶土培, 建立了水稻()根干重与根长、根体积及根表面积的幂函数方程。根系生物量与根系生态指标所处环境相同, 因此直接建立根系生物量与根系生态指标的模拟方程, 可忽略外界环境对根系生物量的影响[15-16], 有助于提高根系生物量预测的准确性。
玉米()作为重要的粮食作物, 种植范围广泛, 其庞大的根系系统在土壤碳循环方面起着重要作用。随着根系观测技术的进步, 玉米根系在形态建成、生长与分布规律等方面的研究也日渐深入[17-18]。微根区管法已经应用于农田生态系统, 对玉米根系的空间分布及生长特征都有了进一步的研究[19-20]。微根区管法可对根的分枝、根径、根长等进行长时间定量检测, 然而无法从图像中直接测得根系生物量、根的死亡和分解[21]。如果能建立根系生物量与根系生态指标的模拟方程, 结合微根区管法可以实现根系生物量的实时观测, 即可弥补根系生物量观测难, 无法省时省力地研究其动态变化的缺陷。基于此目的, 本文利用挖土法和根系扫描系统, 试图建立玉米根系生物量与根系生态指标之间的模拟方程, 这在全球变化的基础研究中具有重要意义。
1 材料与方法
1.1 研究区概况
试验于2016年9月26日在辽宁省锦州市农业气象试验站进行(41°49′N, 121°12′E)。该区属温带季风气候, 年平均气温7.8~9.0 ℃, 年极端最高气温41.8 ℃, 年极端最低气温-31.3 ℃, 年无霜期144~180 d, 年平均降水量540~640 mm。土壤为典型棕壤。种植作物为玉米, 每年1茬, 4月底或5月初种植, 9月底成熟, 10月初收获。整个生长季无灌溉, 播种前条翻, 种植前一次性施氮肥, 施肥量为300 kg∙hm-2。
1.2 根系样品的采集与测定
在玉米成熟期用挖土法取样, 取样位置以1棵玉米植株为中心, 取样样方为50 cm×50 cm, 每10 cm为一层, 取样深度为150 cm。3次重复。分层冲洗, 根系在75 ℃恒温条件下烘干48 h。每层的根系用精度0.000 1的天平称重。
EPSON扫描仪扫描随机选取烘干后的玉米根系, 获取根系扫描图片。根系扫描结束后, 利用精度0.000 1 g天平称重, 得到每张扫描图片根系的干重。玉米根系扫描图片用根系分析系统(WinRHIZO)进行分析, 获取根系的总长度、平均直径、总根表面积、分枝数、交叉数和根尖数等生态指标。
EPSON扫描仪扫描玉米根系过程中发现根系有重叠现象, 不利于准确分析根系扫描图片。扫描玉米根系的目的是获取根系生态指标, 以期建立根系生物量的模拟方程, 而根系的某些生态指标不影响根系生物量的模拟, 如根尖数、分枝数和交叉数, 因此, 人为剥离根系的主根和侧根, 使其无重叠地分布于扫描仪中, 这样更有利于更精确地获取根系生态指标, 有助于提高根系生物量模型模拟的精度。
1.3 建模方法
利用随机扫描及称重获取的玉米根系生物量进行建模, 用于建模的数据共分149组, 分组标准参考文献[22], 80%数据组用于构建根系生物量模型, 20%数据组用于根系生物量模型的验证。结合玉米根系生物量与生态指标的散点图及前人相关研究[22-23], 分别选取根径()、根长()及二者的组合变量(2)建立根系生物量的回归模型。


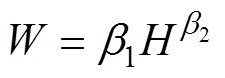
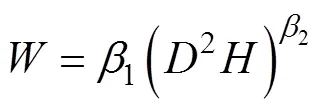
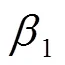
根系生物量模型的验证选取决定系数(2)、平均绝对误差(MAE)、估计值的标准误差(SEE)、平均预估误差(MPE)4项统计指标[9]。其计算公式如下:


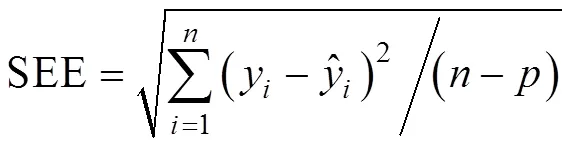

2 结果与分析
2.1 玉米根系生物量的垂直分布特征
玉米根系生物量为玉米根系在土壤体积为25 000 cm3(长×宽×高)的质量(g)。玉米根系生物量的垂直分布总体随土壤深度的增加而减少(图1)。玉米根系生物量主要集中于0~30 cm土壤深度, 总生物量为164.07 g, 占垂直分布总量的94.44%, 与土壤深层的根系生物量存在显著差异性(<0.05)。30~40 cm土层较20~30 cm土层, 玉米根系生物量骤降, 降幅达88.54%。50 cm以下各土层根系生物量降幅相对较平缓, 变异系数为60.73%, 其中40~50 cm土层根系生物量最高, 为1.36 g, 140~150 cm土层根系生物量最低, 为0.24 g。
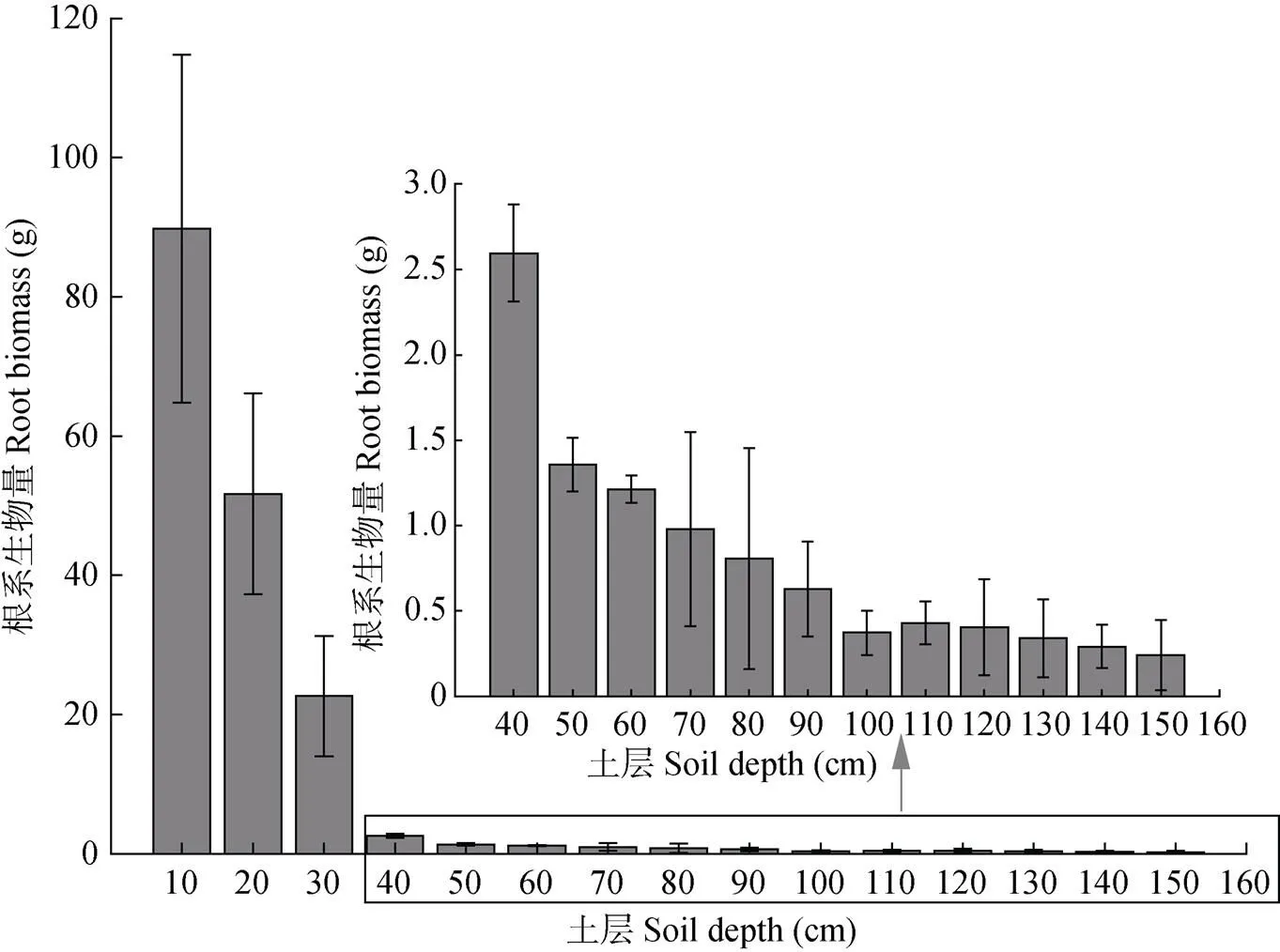
图1 玉米根系生物量的垂直分布
根系生物量测量的土壤体积为25 000 cm3。The soil volume for measurement of root biomass is 25 000 cm3.
2.2 普通最小二乘法建立根系生物量模型
根据选取的模型, 采用普通最小二乘法计算模型的参数, 结果见表1。玉米根系生物量以根径为参数建立的指数函数与幂函数,2分别为0.10和0.12, 说明玉米根系生物量受根径的影响较小, 无法满足构建根系生物量模型的基本要求。玉米根系生物量以根长()或根长与根径的结合变量(2)构建模型,2分别为0.91和0.95, 可见玉米根系生物量模型增加根长为自变量后模拟效果较好。
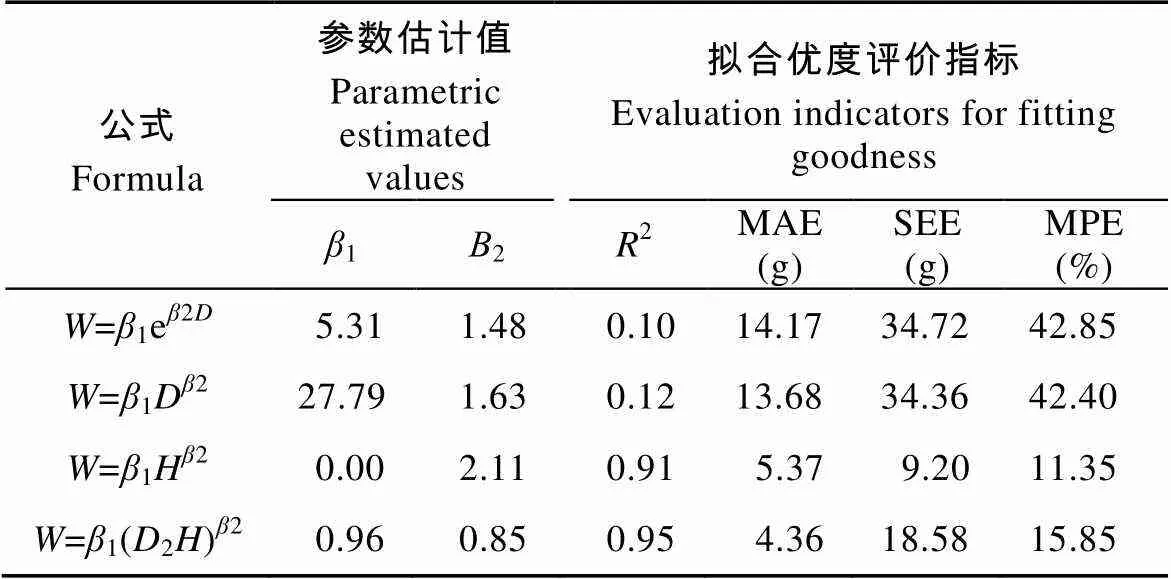
表1 玉米根系生物量回归模型及其拟合优度评价(基于生物量乘以100后计算)
: 根系生物量;: 根径;: 根长;2: 决定系数; MAE: 平均绝对误差; SEE: 估计值的标准误差; MPE: 平均预估误差。: root biomass;: root diameter;: root length;2: coefficient of determination; MAE: mean absolute error; SEE: standard error of estimate; MPE: mean prediction error.
玉米根系生物量模型拟合优度评价指标比较发现, 由根长建立的幂函数除MAE外, SEE和MPE均最小, 分别为9.20 g和11.35%; 由2建立的幂函数MAE最小, 为4.36 g。结合模型的决定系数和拟合优度评价指标, 发现由根长及根长与根径的结合变量(2)建立的回归模型都能较好地模拟根系生物量。进一步对回归模型进行残差分析发现, 随着解释变量的增大, 4个模型的生物量残差均随之增大, 基本呈喇叭形状(图2), 说明4个玉米根系生物量模型均有异方差存在。在回归理论中已经证明模型中存在异方差性, 普通最小二乘法不再适用, 模型的估计不再具有最小方差的特性, 系数的显著性检验也失去意义[24]。尽管或2模拟的玉米根系生物量模型2较高, MAE、SEE和MPE较低, 但因为异方差的存在, 模型依然不能稳定地或相对准确地预测根系生物量。
2.3 消除异方差后建立根系生物量模型及验证
回归模型参数的估计采用普通最小二乘法, 该方法要求模型中的误差项必须满足零数学期望、独立和等方差[24], 因此玉米根系生物量模型必须采用一定的方法消除异方差。回归模型中消除异方差, 通常采用加权法或对数据进行转换来消除[23-24]。本研究中生物量模型采用对数转换的方法, 将上述4个非线性模型转化为相应的线性模型, 然后利用普通最小二乘法计算转化为线性模型的参数, 最后将线性模型参数估计值还原为非线性模型的参数估计值。玉米根系生物量模型经对数转换后, 玉米根系生物量对数值的残差基本上随机地分布在零水平线上下两侧, 数据的异方差问题明显改善(图3)。
从表2可知, 采用对数转换消除异方差后, 进一步检验玉米根系生物量模型, 发现由根径建立的指数函数或幂函数的显著性水平提高,2分别为0.32和0.35, 而根长构建的生物量模型显著水平明显下降,2为0.20, 根长和根径的结合变量(2)构建的根系生物量模型2为0.90, 显著性水平稍有下降。进一步对4个回归模型的拟合效果进行检验发现, 由根长和根径的结合变量(2)构建的根系生物量模型评价指标均最小, MAE、SEE和MPE分别为4.38 g、18.68 g和16.09%。结合模拟的显著性分析, 经对数转换消除异方差后由根长和根径的结合变量(2)构建的模型是模拟玉米根系生物量的最优模型。选取此模型, 利用29组根长和根径数据计算模拟值, 将模拟值和实测值作相关性分析, 发现模拟值和实测值较接近(图4), 相关系数为0.92, 达极显著(<0.01), 这进一步验证了此模型可以模拟玉米根系生物量。
3 讨论
玉米根系生物量随土壤深度的增加而减少, 主要集中于0~30 cm土层, 这和李潮海等[25]的研究结果基本一致。根系生物量的垂直分布特征应该与根系的构型特点有关。玉米根系是由轴根及发生于其上的侧根组成, 而轴根是支撑整个根系的骨架, 根系生物量主要由轴根的数量决定[26]。玉米根系生物量主要集中于0~30 cm的土壤深度, 可能与轴根较多地分布在0~30 cm土层有关。土壤深度50 cm以下, 玉米根系生物量的降幅比较平缓, 这与其他研究者的结论基本一致[17,27]。研究地点不同, 玉米农田的管理方式也不同, 但玉米根系生物量总体变化趋势一致, 进一步说明根系的构型特点决定了根系生物量在土壤中的分布。
: 根系生物量;: 根径;: 根长;: 残差。: root biomass;: root diameter;: root length;: residual.
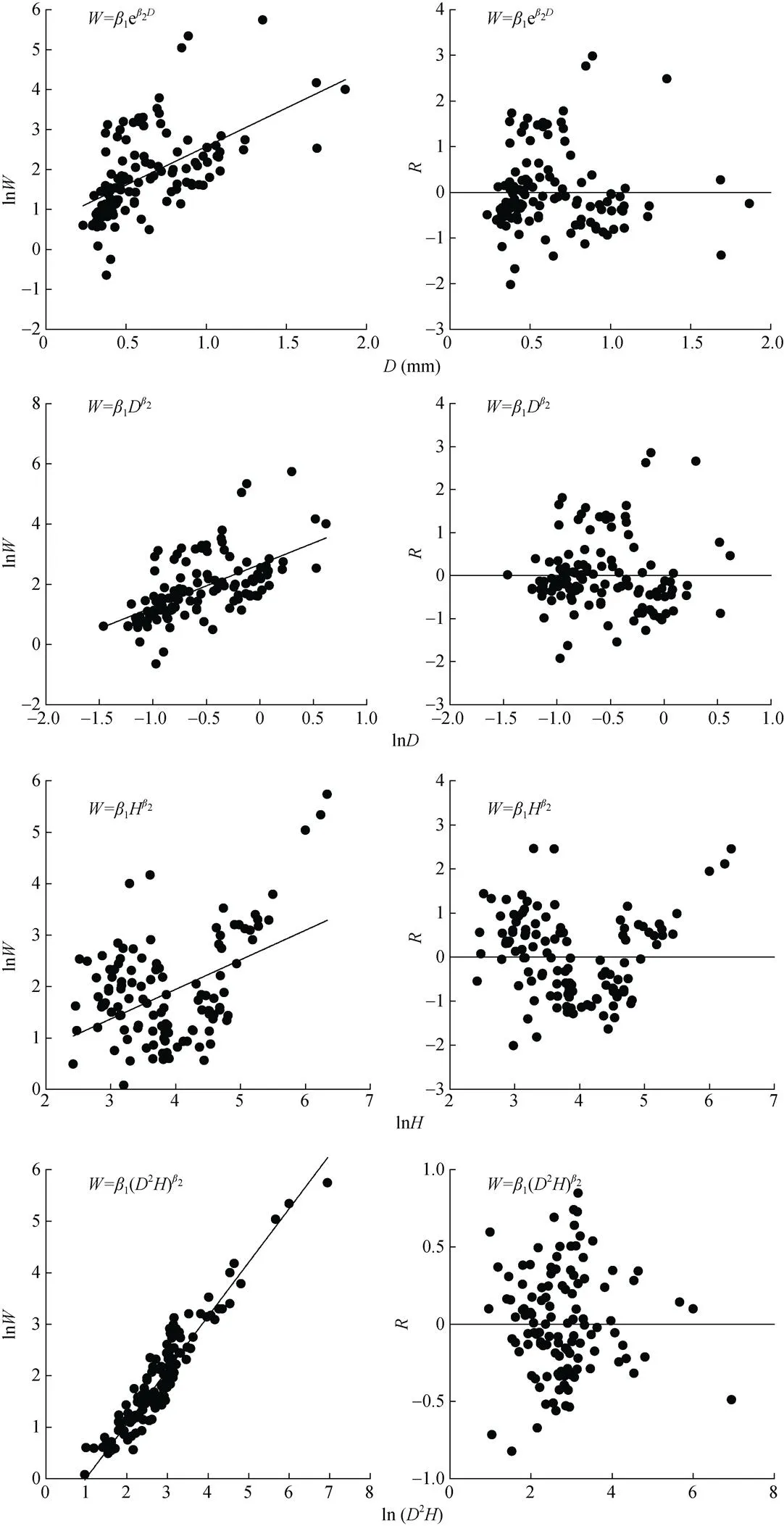
图3 消除异方差后玉米根系生物量模型的构建及残差分析
: 根系生物量;: 根径;: 根长;: 残差。: root biomass;: root diameter;: root length;: residual.
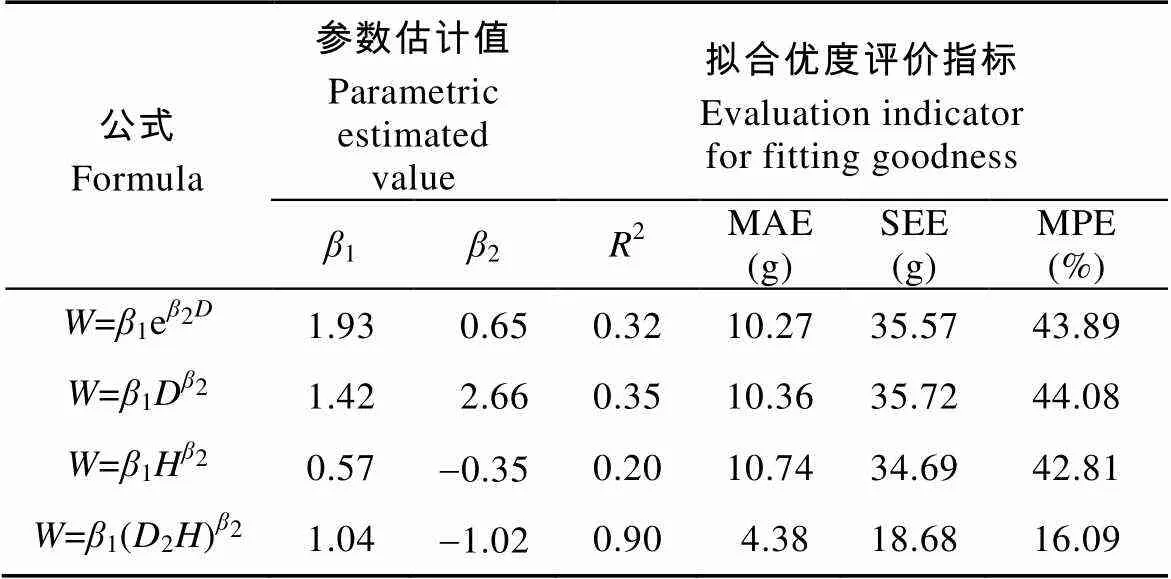
表2 消除异方差后玉米根系生物量回归模型及其拟合优度评价(基于生物量乘以100后计算)
: 根系生物量;: 根径;: 根长;2: 决定系数; MAE: 平均绝对误差; SEE: 估计值的标准误差; MPE: 平均预估误差。: root biamass;: root diameter;: root length;2: coefficient of determination; MAE: mean absolute error; SEE: standard error of estimate; MPE: mean prediction error.
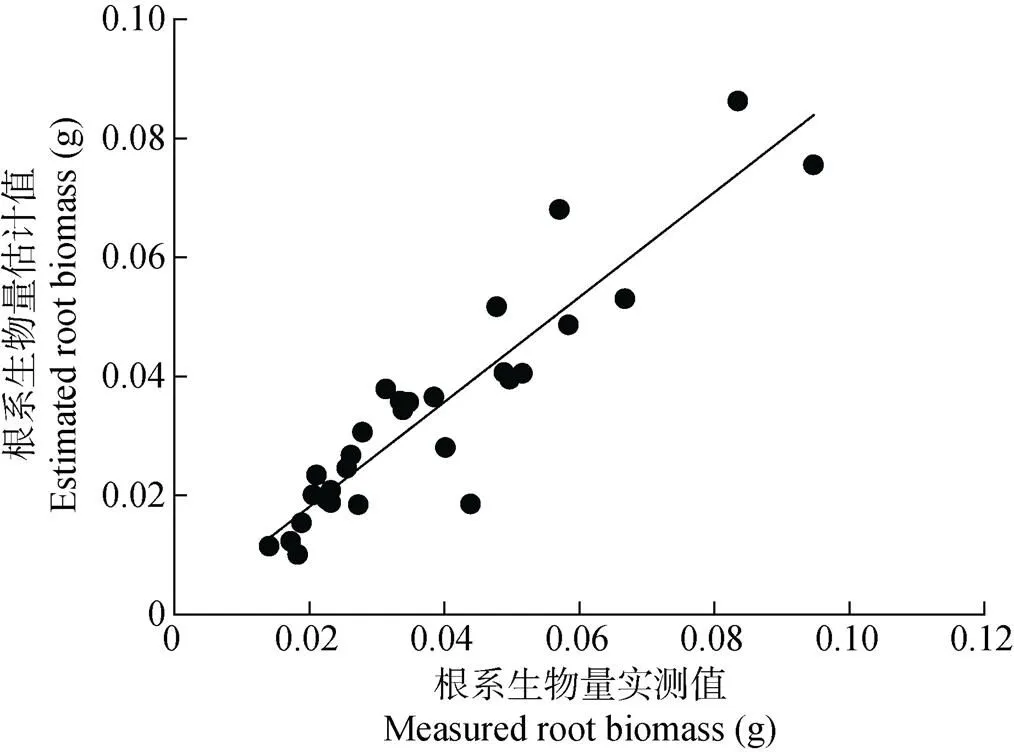
图4 玉米根系生物量模型的实测值和模型估计值
作物根系生物量的模拟多是基于相对综合的机理模型, 更多地是考虑气象要素和生物量分配, 如CERE-MAIZE模型和Rootstimu4.0[16]。农作物相比于森林, 传统的取根法如挖土法、钻土芯法或者室内盆栽法均可以实现根系生物量的直接观测, 所以根系生物量的模拟大多集中于森林生态系统[7-10]。随着根系观测技术微根管法的出现, 直接观测根系生态指标成为可能, 因此利用根系生态指标直接模拟根系生物量成为可能。本研究中, 通过比较模拟生物量常用的模型来模拟玉米根系生物量, 结果发现, 玉米根系生物量模型模拟的最佳参数是根长和根径的组合变量(2)。此模型为二元模型, 是生物量估计中常用的模型[22]。从根系的几何构型来说,2是反映根系体积的参数组合, 与根系生物量关系密切, 此研究结果与杨宪龙等[22]的研究结论一致。
以或做一元模拟时发现, 玉米根系生物量与根长的关系密切, 模拟方程的2达0.91, 而与根径的模拟方程无论是幂函数还是指数函数,2均不足0.15。玉米根系是须根系, 包括初生根、次生根和气生根[17]。玉米根系根径数量级相对较小, 而根长数量级则较大, 如刘晶淼等[19]研究发现玉米根系长度总量可达8.288 km∙m-2。另外, 在玉米不同发育期根系长度差异性较大, 灌浆盛期根系总长度大约是拔节期根系总长度的4~5倍[28], 因此根长较根径对玉米根系构型的影响更高, 从而对根系生物量的影响也较大。黄建辉等[21]也在森林生态系统根系生物量研究进展中提到根系影像分析系统不能直接测定根系生物量, 只能通过根的长度来估计, 这从侧面进一步说明根系生物量与根长关系密切。
生物量建模过程中, 往往存在异方差性, 会导致采用普通最小二乘法进行回归所得估计值有偏, 参数值和有效区间失真[29]。本研究采用普通最小二乘法建模后随着解释变量的增大, 4个模型的生物量残差均随之增大, 基本呈喇叭形状, 说明4个根系生物量确实存在异方差问题。异方差问题的存在会降低模型估算的精度, 因此必须消除。4个生物量模型经对数转换后, 异方差问题明显改善, 然而模型的优劣还需要有一系列的评价指标进行评价。由根径建立的指数函数或幂函数消除异方差后,2明显提高, 然而还远不能达到可以很好地模拟根生物量的标准,2分别为0.32和0.35。根长的指数函数消除异方差后,2明显下降, 为0.20, 说明对数转换不适合此模型的模拟, 对其异方差的消除方法还需要进一步的探讨。2组合变量经对数转换后模型的决定系数为0.90, 其中MAP值为16.09%, 说明2组合变量模型消除异方差后, 平均预估精度达84%左右。此模型的其他评价指标较其他模型也最低, 说明此模型是模拟根系生物量的最优模型。进一步验证此模型, 根系生物量的估计值和实测值相关系数达0.92, 进一步说明选取的最优模型适合模拟玉米根系生物量。此研究中玉米根系通过挖土法获取, 该方法在洗根过程中会损失部分根系, 特别是须根部分, 然而, 本研究中模型构建的数据来源是基于洗根后经扫描和称重得到相应的参数, 因此挖土法损失的须根不影响模型的构建。然而, 玉米根系的须根部分往往是代谢较活跃部分, 因此深入研究不同根径水平下根系生物量模型的构建, 可能更有利于提高陆面过程模型的模拟精度。
4 结论
玉米根系生物量的垂直分布总体随土壤深度的增加而减少, 主要集中在0~30 cm土层, 50 cm以下土层根系生物量变化较小。
普通最小二乘法建立根系生物量模型中, 以根径为自变量建立的指数函数与幂函数模拟精度较低,2分别为0.10和0.12。增加根长作为自变量构建的生物量模型显著地提高了模拟精度,2达0.91以上。普通最小二乘法建立的根系生物量模型均存在异方差问题, 模型不能稳定地或相对准确地预测根系生物量。
采用对数转换可消除模型的异方差问题。经比较发现选取根长和根径的组合变量(2)建立的玉米根系生物量模型具有较好的模拟效果和预测精度, 模拟方程的2为0.90, 评价指标MAE、SEE和MPE分别为4.38 g、18.68 g和16.09%。进一步验证此模型, 发现模拟值和实测值的相关系数为0.92, 达极显著性(<0.01), 说明此模型可以用于模拟玉米根系生物量。
[1] 陈红, 冯云, 周建梅, 等. 植物根系生物学研究进展[J]. 世界林业研究, 2013, 26(5): 25–29 CHEN H, FENG Y, ZHOU J M, et al. Research advance of plant root biology[J]. World Forestry Research,2013, 26(5): 25–29
[2] 罗东辉, 夏婧, 袁婧薇, 等. 我国西南山地喀斯特植被的根系生物量初探[J]. 植物生态学报, 2010, 34(5): 611–618 LUO D H, XIA J, YUAN J W, et al. Root biomass of karst vegetation in a mountainous area of southwestern China[J]. Chinese Journal of Plant Ecology, 2010, 34(5): 611–618
[3] 郭京衡, 李尝君, 曾凡江, 等. 2种荒漠植物根系生物量分布与土壤水分、养分的关系[J]. 干旱区研究, 2016, 33(1): 166–171 GUO J H, LI C J, ZENG F J, et al. Relationship between root biomass distribution and soil moisture, nutrient for two desert plant species[J]. Arid Zone Research, 2016, 33(1): 166–171
[4] 张金凤, 徐雨晴. 水氮添加对内蒙古多伦县退耕还草地生物量、生产力及其分配的影响[J]. 中国生态农业学报, 2016, 24(2): 192–200 ZHANG J F, XU Y Q. Responses of plant biomass and net primary production to nitrogen fertilization and increased precipitation in re-grassed croplands in Duolun County of Inner Mongolia, China[J]. Chinese Journal of Eco-Agriculture, 2016, 24(2): 192–200
[5] 韩畅, 宋敏, 杜虎, 等. 广西不同林龄杉木、马尾松人工林根系生物量及碳储量特征[J]. 生态学报, 2017, 37(7): 2282–2289 HAN C, SONG M, DU H, et al. Biomass and carbon storage in roots ofandplantations at different stand ages in Guangxi[J]. Acta Ecologica Sinica, 2017, 37(7): 2282–2289
[6] 周本智, 张守攻, 傅懋毅. 植物根系研究新技术Minirhizotron的起源、发展和应用[J]. 生态学杂志, 2007, 26(2): 253–260 ZHOU B Z, ZHANG S G, FU M Y. Minirhizotron, a new technique for plant root system research: Its invention, development and application[J]. Chinese Journal of Ecology, 2007, 26(2): 253–260
[7] ZIANIS D, MUUKKONEN P, MÄKIPÄÄ R, et al. Biomass and stem volume equations for tree species in Europe[J]. Silva Fennica Monographs, 2005, 4: 1–63
[8] 崔喜红, 陈晋, 沈金松, 等. 基于探地雷达的树木根径估算模型及根生物量估算新方法[J]. 中国科学: 地球科学, 2011, 41(2): 243–252 CUI X H, CHEN J, SHEN J S, et al. Modeling tree root diameter and biomass by ground-penetrating radar[J]. Science China Earth Sciences, 2011, 41(2): 243–252
[9] 曾伟生, 唐守正. 东北落叶松和南方马尾松地下生物量模型研建[J]. 北京林业大学学报, 2011, 33(2): 1–6 ZENG W S, TANG S Z. Establishment of below-ground biomass equations for larch in northeastern and masson pine in southern China[J]. Journal of Beijing Forestry University, 2011, 33(2): 1–6
[10] 肖义发, 欧光龙, 王俊峰, 等. 思茅松单木根系生物量的估算模型[J]. 东北林业大学学报, 2014, 42(1): 57–60 XIAO Y F, OU G L, WANG J F, et al. Biomass estimation models of individual root forvar.[J]. Journal of Northeast Forestry University, 2014, 42(1): 57–60
[11] LITTON C M, RYAN M G, TINKER D B, et al. Belowground and aboveground biomass in young postfire lodgepole pine forests of contrasting tree density[J]. Canadian Journal of Forest Research, 2003, 33(2): 351–363
[12] BOLTE A, RAHMANN T, KUHR M, et al. Relationships between tree dimension and coarse root biomass in mixed stands of European beech (L.) and Norway spruce (L. Karst.)[J]. Plant and Soil, 2004, 264(1/2): 1–11
[13] JOHANSSON T. Biomass production and allometric above- and below-ground relations for young birch stands planted at four spacings on abandoned farmland[J]. Forestry, 2007, 80(1): 41–52
[14] 刘永霞, 岳延滨, 刘岩, 等. 基于生物量的水稻根系生长动态模型[J]. 江苏农业学报, 2011, 27(4): 704–709 LIU Y X, YUE Y B, LIU Y, et al. Biomass-based dynamic model for rice root system[J]. JiangsuJournal of Agricultural Sciences, 2011, 27(4): 704–709
[15] 王空军, 郑洪建, 刘开昌, 等. 我国玉米品种更替过程中根系时空分布特征的演变[J]. 植物生态学报, 2001, 25(4): 472–475 WANG K J, ZHENG H J, LIU K C, et al. Evolution of maize root distribution in space-time during maize varieties replacing in China[J]. Acta Phytoecologica Sinica, 2001, 25(4): 472–475
[16] 许常华, 李荣平, 祁红彦, 等. 植物根系模拟进展[J]. 中国农学通报, 2014, 30(18): 18–22 XU C H, LI R P, QI H Y, et al. Study progress of plant root model[J]. Chinese Agricultural Science Bulletin, 2014, 30(18): 18–22
[17] 管建慧, 刘克礼, 郭新宇. 玉米根系构型的研究进展[J]. 玉米科学, 2006, 14(6): 162–166 GUAN J H, LIU K L, GUO X Y. Advances of research on maize root system architecture[J]. Journal of Maize Sciences, 2006, 14(6): 162–166
[18] 刘蕾. 中国玉米根系生物量及空间分布特征[D]. 北京: 中国农业大学, 2016 LIU L. Biomass estimation and spatial distribution of maize root in China[D]. Beijing: China Agricultural University, 2016
[19] 刘晶淼, 安顺清, 廖荣伟, 等. 玉米根系在土壤剖面中的分布研究[J]. 中国生态农业学报, 2009, 17(3): 517–521 LIU J M, AN S Q, LIAO R W, et al. Temporal variation and spatial distribution of the root system of corn in a soil profile[J]. Chinese Journal of Eco-Agriculture, 2009, 17(3): 517–521
[20] 廖荣伟, 刘晶淼, 安顺清, 等. 基于微根管技术的玉米根系生长监测[J]. 农业工程学报, 2010, 26(10): 156–161 LIAO R W, LIU J M, AN S Q, et al. Monitor of corn root growth in soil based on minirhizotron technique[J]. Transactions of the CSAE, 2010, 26(10): 156–161
[21] 黄建辉, 韩兴国, 陈灵芝. 森林生态系统根系生物量研究进展[J]. 生态学报, 1999, 19(2): 270–277 HUANG J H, HAN X G, CHEN L Z. Advances in the research of (fine) root biomass in forest ecosystems[J]. Acta Ecologica Sinica, 1999, 19(2): 270–277
[22] 杨宪龙, 魏孝荣, 邵明安. 黄土高原北部典型灌丛枝条生物量估算模型[J]. 应用生态学报, 2016, 27(10): 3164–3172 YANG X L, WEI X R, SHAO M A. Stem biomass estimation models for dominant shrubs on the northern Loess Plateau of China[J]. Chinese Journal of Applied Ecology, 2016, 27(10): 3164–3172
[23] 曾伟生. 3种异速生长方程对生物量建模的对比分析[J]. 中南林业调查规划, 2014, 33(1): 1–3 ZENG W S. Comparison of three allometric equations for biomass modeling[J]. Central South Forest Inventory and Planning, 2014, 33(1): 1–3
[24] 张会儒, 唐守正, 胥辉. 关于生物量模型中的异方差问题[J]. 林业资源管理, 1999, (1): 46–49 ZHANG H R, TANG S Z, XU H. The problem of heteroscedasticity in the biomass model[J]. Forest Resources Management, 1999, (1): 46–49
[25] 李潮海, 李胜利, 王群, 等. 不同质地土壤对玉米根系生长动态的影响[J]. 中国农业科学, 2004, 37(9): 1334–1340 LI C H, LI S L, WANG Q, et al. Effect of different textural soils on root dynamic growth in corn[J]. Scientia Agricultura Sinica, 2004, 37(9): 1334–1340
[26] 李春俭, 彭云峰, 牛君仿, 等. 土壤中的玉米根系生长及其研究应注意的问题[J]. 植物营养与肥料学报, 2010, 16(1): 225–231 LI C J, PENG Y F, NIU J F, et al. Real maize roots in the soil and issues should be considered by study[J]. Plant Nutrition and Fertilizer Science, 2010, 161(1): 225–231
[27] 赵江, 张怡明, 牛兴奎, 等. 不同密度条件玉米根系性状在不同土层中的分布研究[J]. 华北农学报, 2011, 26(S1): 99–103 ZHAO J, ZHANG Y M, NIU X K, et al. Studies on the distribution of maize root characteristics at different soil layers and densities[J]. Acta Agriculturae Boreali-Sinica, 2011, 26(S1): 99–103
[28] 张玉, 秦华东, 伍龙梅, 等. 玉米根系空间分布特性的数学模拟及应用[J]. 玉米科学, 2015, 23(4): 92–97 ZHANG Y, QIN H D, WU L M, et al. Mathematical simulation of maize root spatial distribution and its application[J]. Journal of Maize Sciences, 2015, 23(4): 92–97
[29] 杨嘉龙, 肖生苓. 异方差对生物量模型构建的影响[J]. 森林工程, 2014, 30(2): 25–28 YANG J L, XIAO S L. The influence of heteroscedasticity on the establishment of biomass model[J]. Forest Engineering, 2014, 30(2): 25–28
Modeling root biomass of maize in Northeast China*
LYU Guohong, XIE Yanbing, WEN Rihong, WANG Xiaoying**, JIA Qingyu
(Shenyang Institute of Atmospheric Environment, China Meteorological Administration, Shenyang 110166, China)
It is of great significance to explore root biomass and establish a universal root biomass model for the monitoring and evaluation of the ecosystem biomass. In order to get the real-time information of a root system, the biomass and ecological indexes of maize root system were collected using soil sampling method and root scanning systems at the agricultural meteorological experiment station in Jinzhou City, Liaoning Province in September 2016. The vertical distribution characteristics of root biomass of maize were analyzed and simulation equations were established based on the relationships between root biomass and root ecological indexes. The results showed that the maize root biomass decreased with increase of soil depth. The root biomass of maize mainly concentrated at the soil depth from 0 cm to 30 cm, which accounted for 94.44% of the total root biomass. The simulation accuracy of exponential and power functions of root biomass with root diameter as independent variable established by ordinary least square method was low, and2was 0.10 and 0.12, respectively. The biomass model constructed by adding root length as an independent variable significantly improved the simulation accuracy, with2reaching above 0.91. There was heteroscedasticity issue in the models of root biomass established by the ordinary least square method inducing less stable and inaccurate prediction results. This issue could be eliminated by using logarithmic transformation. The biomass model of maize root system with root length and root dimeter together as variables (2) had a better simulation effect and a good prediction accuracy, with the coefficient of determination (2) as 0.90 and the evaluation indexes MAE, SEE and MPE as 4.38 g, 18.68 g and 16.09%, respectively. The correlation coefficient between simulated and measured values was 0.92 (< 0.01), indicating that this model could be used to simulate the biomass of maize root system in Northeast China. The study results indicated that the difficulty in observing root biomass in real time could be resolved by using root biomass model combined with the minirhizotron method.
Maize; Root biomass; Root length; Root diameter
, E-mail: wangxy_0917@qq.com
Jan. 25, 2018;
Jul. 5, 2018
10.13930/j.cnki.cjea.180115
S513
A
2096-6237(2019)04-0572-09
王笑影, 主要研究方向为生态气象。E-mail: wangxy_0917@qq.com
吕国红, 主要研究方向为生态气象。
2018-01-25
2018-07-05
* This research was supported by the National Natural Science Foundation of China (31501215) and the Special Scientific Research Fund of Meteorology in Public Welfare of China (2016SYIAEZD).
* 国家自然科学基金项目(31501215)和公益性行业(气象)科研专项基金(2016SYIAEZD)资助
吕国红, 谢艳兵, 温日红, 王笑影, 贾庆宇. 东北玉米根系生物量模型的构建[J]. 中国生态农业学报(中英文), 2019, 27(4): 572-580
LYU G H, XIE Y B, WEN R H, WANG X Y, JIA Q Y. Modeling root biomass of maize in Northeast China[J]. Chinese Journal of Eco-Agriculture, 2019, 27(4): 572-580
