硒砂瓜连作对土壤真菌群落结构的影响*
2019-04-13冯翠娥岳思君简阿妮陈丽萍苏建宇
冯翠娥, 岳思君, 简阿妮, 陈丽萍, 郭 洋, 郑 蕊, 苏建宇
硒砂瓜连作对土壤真菌群落结构的影响*
冯翠娥, 岳思君**, 简阿妮, 陈丽萍, 郭 洋, 郑 蕊, 苏建宇**
(宁夏大学生命科学学院/西部特色生物资源保护与利用教育部重点实验室 银川 750021)
硒砂瓜是宁夏地区重要的经济作物, 其连作严重影响硒砂瓜产量和品质。目前硒砂瓜连作对土壤真菌群落结构的影响尚不清楚。本研究采用Illumina MiSeq高通量测序技术, 探讨硒砂瓜连作对土壤真菌群落结构的影响。研究发现, 硒砂瓜连作土壤中真菌群落多样性指数和丰富度指数随连作年限的增加先上升后下降。供试土壤样本中共检测到真菌8门、25纲、244属, 其中子囊菌门(Ascomycota)、接合菌门(Zygomycota)是优势菌门, 占90%以上。与对照相比, 连作30年土壤中子囊菌门丰度下降32.51%, 接合菌门丰度上升29.89%。供试土壤中真菌主要的优势属为被孢霉属()、绿僵菌属()、假霉样真菌属()、镰刀菌属()和青霉属()。与对照相比, 连作5年土壤中假霉样真菌属丰度增加45.81%, 连作10年土壤中镰刀菌属丰度增加26.74%, 连作15年土壤中绿僵菌属下降26.83%, 连作20年土壤中青霉属增加29.68%, 连作25年土壤中绿僵菌属减少18.30%, 连作30年土壤中被孢霉属丰度上升29.89%。土壤理化性质与硒砂瓜连作年限间无显著相关性, 而与土壤真菌群落结构存在显著的相关性。土壤全磷、碱解氮、有效磷含量是影响土壤真菌群落最主要的因素。研究结果表明, 导致硒砂瓜连作障碍的主要原因不是土壤理化性质变化, 而是土壤真菌群落结构的改变。研究结果可为硒砂瓜土传病害的生物防治提供参考依据。
连作; 土壤真菌; 群落结构; Illumina高通量测序; 土壤理化性质; 硒砂瓜
西瓜[(Thunb.) Matsum. et Nakai]为葫芦科(Cucurbitaceae)一年生蔓生藤本植物, 在宁夏中卫市环香山地区是一种重要的经济作物。该地区主要采用一种特有的保温、保水、保墒的旱作农业种植模式——沙地上压砂石进行农业耕作, 种出的西瓜富含硒等微量元素, 因此得名“硒砂瓜”。近年来, 由于西瓜枯萎病导致西瓜连作障碍严重, 严重影响硒砂瓜产量和品质。因此, 采用合理有效的微生物学方法进行硒砂瓜连作障碍的防治具有重要意义。
真菌是土壤微生物的重要组成部分之一, 对土壤中生物化学物质的循环及土传病害的监测等起着不可替代的驱动作用[1], 同时也会引起70%~80%的植物病害[2]。作物在长期种植过程中, 由于管理与种植方式相同, 为土壤微生物提供了相同的微生态环境, 定向地影响土壤微生物的生长繁殖, 使得土壤微生物多样性降低, 土传病害增加, 土壤微生物活力下降[3], 微生物结构组成失衡[4]。有研究表明, 作物长期种植所导致的土壤真菌群落结构改变是导致土壤连作障碍的主要原因[5-6]。西瓜枯萎病是西瓜连作过程中最为严重的真菌病害, 其病原菌主要为镰刀菌属的尖孢镰刀菌西瓜专化型(f. sp.)[7]、腐皮镰刀菌葫芦专化型(f. sp.)[8]等, 会导致西瓜产量降低和品质下降[9]。
硒砂瓜连作严重影响宁夏地区硒砂瓜产量及品质。目前, 对于硒砂瓜的研究主要集中在产业的发展以及机械化生产等方面[10-11], 而对硒砂瓜连作土壤微生物多样性研究少有报道。近年来, 连作障碍是农业发展中研究的热点问题之一[12], 但对于宁夏中卫市环香山地区硒砂瓜的连作障碍鲜有研究。研究其连作条件下不同连作年限土壤真菌群落结构, 可为进一步研究连作土壤微生物多样性、减少有害真菌的积累、改善硒砂瓜品质等提供理论基础, 对研究硒砂瓜土传病害的生物防治及更好地发展硒砂瓜产业具有重要意义和实践价值。
1 材料与方法
1.1 研究地概况及试验设计
试验地点位于宁夏中卫市沙坡头区兴仁镇(104°17′~106°10′E、36°06′~37°50′N), 海拔1 100~2 955 m, 属温带干旱气候, 年平均气温8.8 ℃, 年降水量179.6 mm, 年蒸发量1 829.6 mm, 农业生产对自然降水依赖性很强。研究地表层土为灰钙土, 地表有大量砂石覆盖, 硒含量为0.003 2 mg∙kg-1。
试验设置7个处理, 分别为连作5年(T5)、10年(T10)、15年(T15)、20年(T20)、25年(T25)、30年(T30)及对照土壤(未种植硒砂瓜土壤, T0), 各处理分别设置3个区, 根据连作时间进行区组设计, 每个区面积为0.33 hm2, 区之间距离较远。硒砂瓜品种为‘金城5号’ 种植时间为4月15—25日。施肥管理采用当地常规措施, 种瓜之前施1次羊粪作为基肥, 在苗期和坐果期追加磷酸二铵肥料。在种植前、苗期、坐果期、膨瓜期分别浇水1次。在果实采收结束后, 除去瓜苗, 以减少病害。其他田间管理措施均相同。
1.2 样本采集
于2017年10月1日在宁夏中卫市沙坡头区兴仁镇硒砂瓜种植大田进行采样。在硒砂瓜采收后期去除表层(0~15 cm)砂石, 取深度为15~25 cm处取土, 每个处理各采集3份土壤, 每份土壤分别在每个区采用五点取样法取样, 除去土壤中的根系、碎石及其他杂物, 混匀成1份土壤装进塑封袋, 放入冰盒带回实验室: 一部分鲜土放入超低温冰箱, 用于微生物多样性研究; 另一部分鲜土放入-4 ℃冰箱保存, 取出其中一部分土壤自然风干后过1 mm筛, 用于测定土壤中全氮、全钾、全磷、碱解氮、速效磷、速效钾、pH、有机质等含量。
1.3 土壤理化性质测定
测定方法: 土壤pH采用电位法(水∶土=2.5∶1), 土壤有机质含量采用重铬酸钾外加热法, 全氮采用凯氏定氮法, 碱解氮采用扩散法, 全磷采用矾钼黄比色法, 有效磷采用钼锑抗比色法, 全钾采用ICP法, 速效钾采用原子吸收分光光度计法。
1.4 土壤微生物群落结构多样性测定
土壤样本送上海美吉生物医药科技有限公司的Illumina Miseq PE300平台(San Diego, CA, USA)进行测序。
1.5 序列分析
原始测序序列使用Trimmomatic[13]软件质控, 使用FLASH软件进行拼接: 去除质控后长度低于50 bp的序列; 去除模糊碱基; 根据重叠碱基overlap 将两端序列进行拼接。使用的UPARSE v7.1软件[14], 对有效数据在97%相似度水平进行操作分类单元OTU聚类; 使用UCHIME软件[15]剔除嵌合体。利用RDP classifier(http://rdp.cme.msu.edu/)[16]对每条序列进行物种分类注释, 比对Unite (Release 6.0 http://unite.ut.ee/index.php)数据库, 设置比对阈值为70%, 并在各个分类水平上统计样本的群落组成。
1.6 数据处理
试验数据采用Microsoft Excel 2016计算, 采用SPSS 19.0进行单因素方差分析(<0.05), 表中数据均采用平均值±标准差表示。
2 结果与分析
2.1 硒砂瓜连作对土壤理化性质的影响
不同连作年限土壤pH均大于8, 为碱性土, 且土壤pH随连作年限的增加呈现先增加后减少趋势, 以T15土壤pH最大; 与T0相比, T5-T25处理土壤有机质、全氮、全磷含量均增加, 在T5中全氮、速效磷、全磷、速效钾、碱解氮含量最高, T15中有机质含量最高。从结果可知, 土壤综合理化性质与硒砂瓜连作年限不存在相关性。因此, 硒砂瓜连作障碍不是由于土壤理化性质的改变所导致的(表1)。
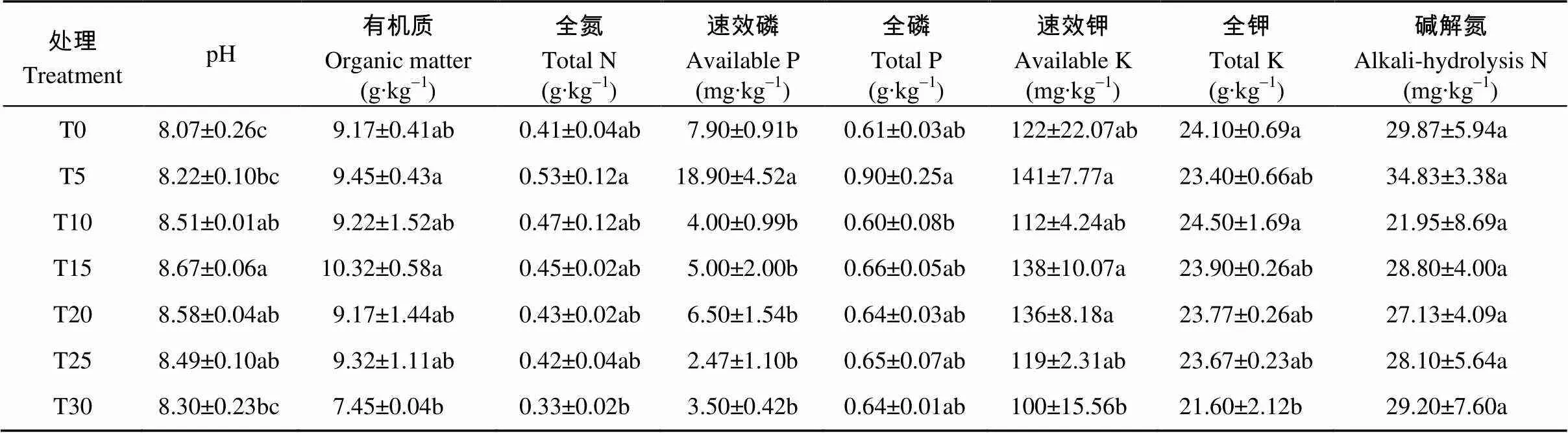
表1 硒砂瓜连作对土壤理化性质的影响
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.
2.2 硒砂瓜连作土壤真菌群落OTU分析
高通量测序结果表明, 从7个样本(3次重复)21份土壤中共获得1 500 251条高质量的真菌基因序列, 平均阅读长度为264.50 bp, 样品中真菌有95%以上的序列长度都分布在221~360 bp, 将这些序列数以97%的序列相似性聚为4 005个不同的OTU。
2.3 硒砂瓜连作土壤真菌群落多样性分析
2.3.1 硒砂瓜土壤样本质量检测
基于Sobs指数的稀释曲线分析表明, 随着样本测序量的增加, 各样本的稀释曲线基本趋向平坦, 获得的序列足以完全捕获真菌的多样性, 说明测序数据合理, 达到测序要求, 更多的数据量只会产生少量新的物种, 能够比较真实地反映土壤样本的真菌群落(图1)。
2.3.2 硒砂瓜连作土壤真菌Alpha多样性指数分析
Alpha多样性可以反映微生物群落的丰富度和多样性。不同连作年限土壤真菌群落的多样性指数如表2所。结果表明, 真菌群落多样性为T15>T10>T20>T0>T30>T5>T25; 物种丰富度为T15>T10>T20>T0>T5> T30>T25。Alpha多样性水平不同, 说明硒砂瓜长期连作会改变土壤真菌群落的组成和分布, T10、T15、T20中的物种多样性指数和丰富度指数均高于T0, 而T25、T30物种多样性指数和丰富度指数低于T0。连作时间对土壤真菌多样性指数和丰富度指数有一定的影响。

图1 硒砂瓜连作土壤样本在OUT水平Sobs指数稀释曲线
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.
2.4 硒砂瓜连作对土壤真菌群落结构的影响
高通量测序结果表明, 所有土壤样本中共有真菌8个门, 25个纲, 244个属, 340个种。样本整体真菌种类相似, 然而在不同分类水平上物种丰富度所占比例不同。在门水平, 物种丰度最高的是子囊菌门(Ascomycota), 其次是接合菌门(Zygomycota)、担子菌门(Basidiomycota)和未分类真菌(unclassified_fungi), 丰度分别为56.38%~88.90%、8.21%~37.10%、4.50%~10.53%、1.49%~4.65%。T0中子囊菌门丰度最高, 随着连作时间的增加, 子囊菌门丰度降低; 接合菌门随连作时间增加, 丰度逐渐增加; 担子菌门在T20中丰度最高(图2)。
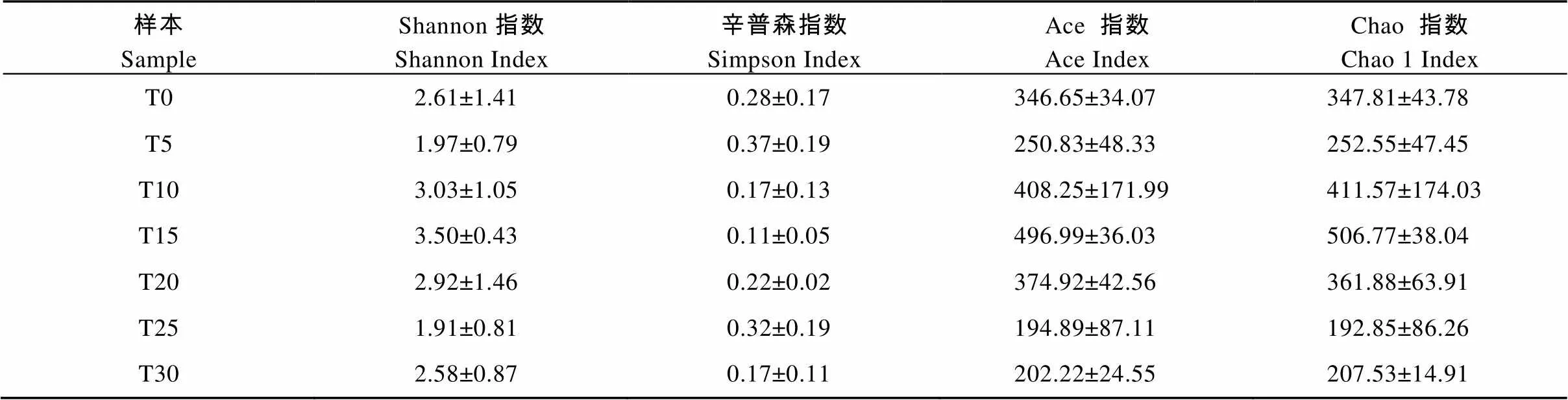
表2 硒砂瓜连作土壤真菌Alpha多样性指数
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.
对不同连作年限土壤真菌群落组成在纲水平进行分析, 丰度前5的物种分别为粪壳菌纲(Sordariomycetes)、norank_p_Zygomycota、散囊菌纲(Eurotiomycetes)、unclassified_p_Ascomycota、座囊菌纲(Dothidemycetes), 其中座囊菌纲在连作T15土壤中丰度最高(9.51%), 散囊菌纲在T20中丰度最高(35.39%), norank_p_Zygomycota在T25中丰度最高(37.10%), 其次是T30(34.29%)。随着连作年限增加,土壤中真菌类群呈逐渐上升的趋势, 其中粪壳菌纲数量逐渐减少, 新出现节担菌纲(Wallemiomycetes)、盘菌纲(Pezizomycetes), 酵母纲(Saccharomycetes)则消失(图3)。
进一步对丰度前30的物种在属水平的Heatmap图分析显示, 丰度前5的属分别为被孢霉属()、绿僵菌属()、假霉样真菌属()、镰刀菌属()、青霉属()。被孢霉属随着连作年限的增加, 丰度逐渐增加; 绿僵菌属在T0中丰度最高, 随连作年限增加呈先增加后下降的趋势; 假霉样真菌属在T5中丰度最高; 镰刀菌属随连作年限增加呈先增加后下降的趋势, 在T10中丰度最高; 青霉属丰度随连作年限增加出现先增加后下降的趋势, 在T20中丰度最高(图4)。
2.5 不同连作年限土壤真菌群落比较
基于Bray-Curtis距离算法对不同连作年限的硒砂瓜土壤真菌群落进行NMDS分析。结果显示, 在T0、T5、T10、T15、T20、T25、T30土壤中部分样品的硒砂瓜土壤真菌群落间距离较近, 组成比较相似, 说明随着连作年限的增加, 硒砂瓜土壤真菌群落组成趋于一致(图5)。
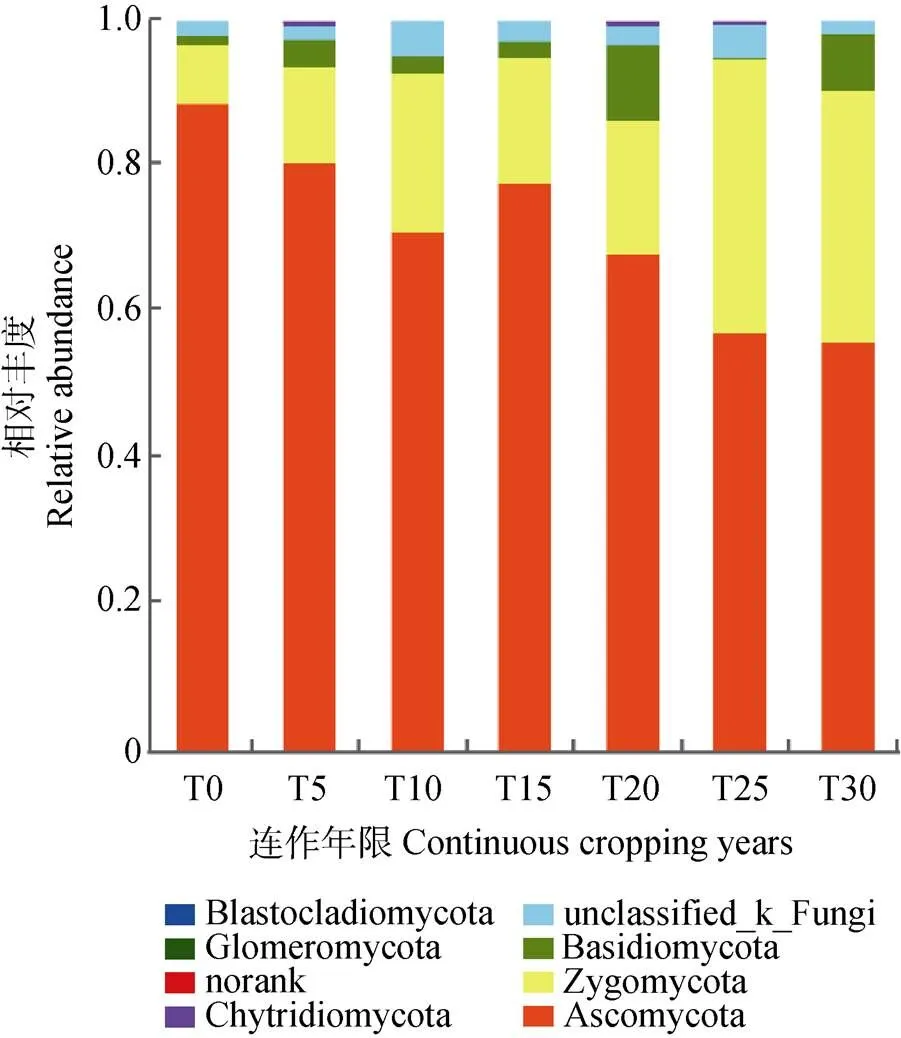
图2 硒砂瓜不同连作年限土壤真菌群落在门水平的相对丰度
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.

图3 硒砂瓜不同连作年限土壤真菌群落在纲水平相对丰度
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.
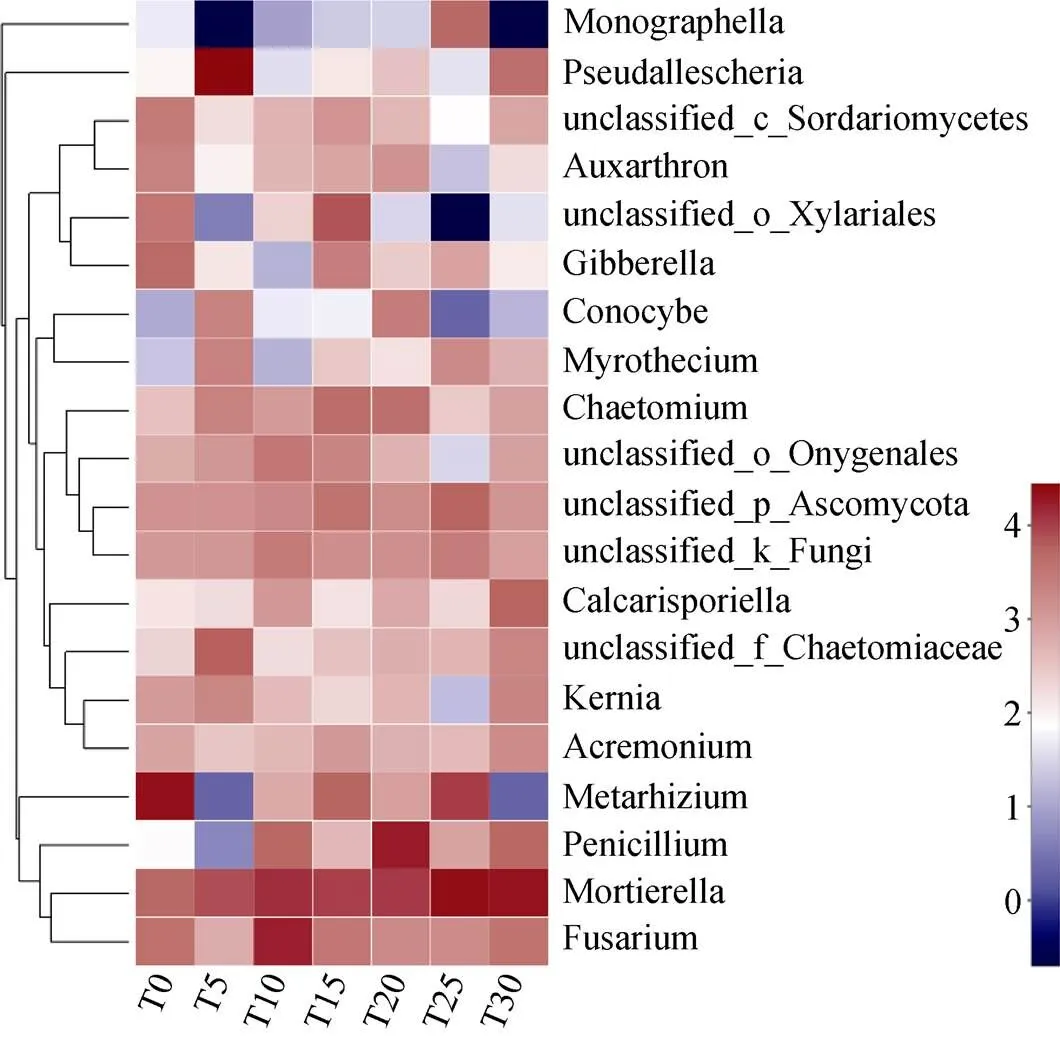
图4 硒砂瓜不同连作年限土壤真菌群落在属水平Heatmap图
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.
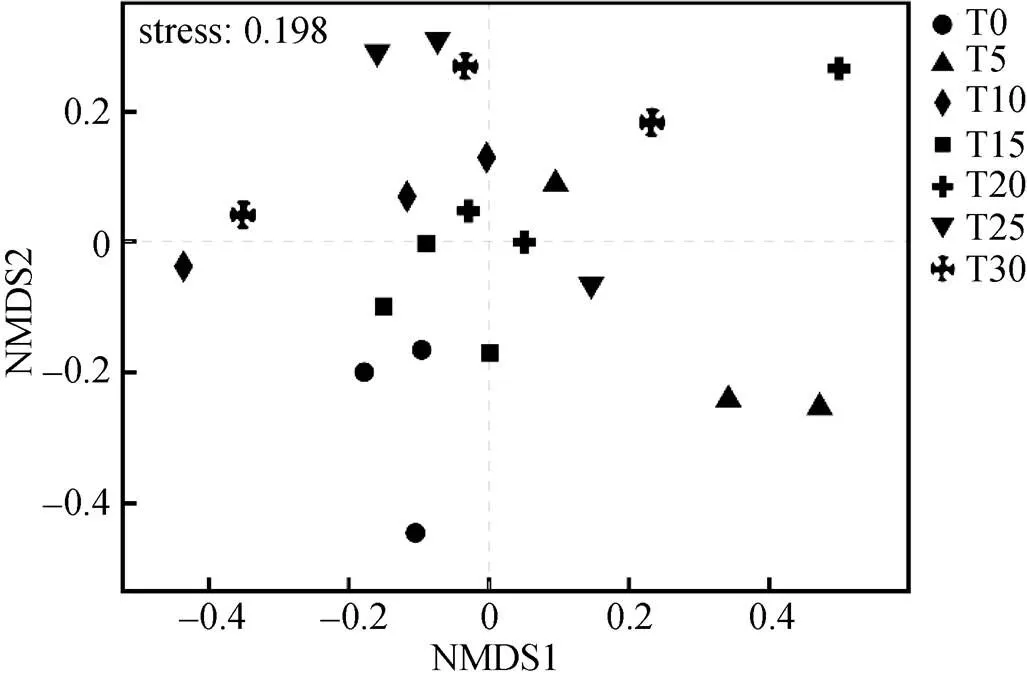
图5 硒砂瓜不同连作年限土壤真菌基于OTU水平的NMDS分析
T0、T5、T10、T15、T20、T25、T30分别指未种植硒砂瓜处理及连作5年、10年、15年、20年、25年和30年处理。T0, T5, T10, T15, T20, T25, and T30 refer to the treatments of non-planting and continuous cropping for 5, 10, 15, 20, 25 and 30 years of selenium melon, respectively.
2.6 硒砂瓜连作土壤真菌群落结构与土壤理化性质的相关性
相关性分析表明, 全磷、碱解氮、速效磷含量是影响土壤真菌群落最主要的因子。被孢霉属丰富度与各土壤理化因子间没有显著的相关性, 绿僵菌属丰富度与全钾含量有显著正相关性(<0.05), 假霉样真菌属丰富度与全磷、速效钾间存在显著(<0.05)和极显著(<0.01)的正相关性, 镰刀菌属丰富度与全磷、碱解氮间存在极显著的负相关性(<0.01), 青霉属丰富度与pH间存在显著正相关性(<0.05), 毛壳属丰富度与速效钾间存在显著正相关性(<0.05)(表3)。

表3 土壤真菌优势菌(属水平)丰度与土壤理化性质的相关性
**< 0.01, *< 0.05.
3 讨论
3.1 硒砂瓜连作对土壤真菌多样性的影响
硒砂瓜是宁夏地区特有的经济作物。有研究表明, 由于长期的连作, 导致土壤肥力下降, 微生物群落多样性降低, 存在潜在的土壤肥力衰退趋势[17]。而在本研究中, 长期连作并未显著改变土壤理化性质。土壤微生物群落多样性能够反映土壤健康状况, 同时微生物群落结构和物种组成多样性的增加能够增加土壤微生态的稳定性。真菌在土壤环境中广泛存在, 对连作土壤中土传病害的发生及作物正常发育有很大影响。本研究中, 长期连作土壤与对照相比, 真菌群落多样性和丰富度指数呈先上升后下降趋势。这一结果与采用磷脂酸生物标记的方法研究不同连作年限怀牛膝(Blume.)土壤微生物多样性结果相似[18]。这可能是由于长期高强度的农业耕作措施破坏了土壤稳定结构, 减小微生物丰度和均匀度, 不利于土壤微生物的代谢, 导致微生物多样性降低。由于硒砂瓜是压砂种植, 对土壤性质或土壤养分的影响较大, 进而使得土壤真菌群落分布不均。
3.2 硒砂瓜连作对真菌群落结构的影响
硒砂瓜土壤中, 在门水平上相对丰度依次为子囊菌门、接合菌门、担子菌门, 与阎海涛等[19]和赵帆等[20]研究结果一致。随着连作年限的增加, 微生物类群的改变和成分的变化会扰乱土壤微生态的平衡, 更严重的会导致植物的死亡[21]。大量研究表明, 微生物多样性会受到连作时间的影响[22]。在本研究中, 随着连作时间的增加子囊菌门丰度逐渐下降。Schoch等[23]研究表明, 子囊菌门在根际土壤有机物的降解中起着重要作用。据此推测子囊菌门可能与土壤有机质降解及土壤肥力下降有关。接合菌门丰度随连作时间的增加而增加, 是引发植物病害和破坏生物多样性的来源之一[24]。因此, 病害的发生可能与子囊菌门及接合菌门丰度的变化有关。
植物的生长与土壤中各种微生物的生理活动密切相关[25]。而真菌在土壤环境中起着重要作用, 同时也是引起农作物病害主要病原菌的来源[26], 其中镰刀菌属是土传病害中主要的病原菌属。本研究中, 被孢霉属丰度随连作时间的增加变化显著, 而被孢霉属是土壤中有益微生物, 具有溶解土壤中磷的功能[27], 并且能够通过提供氮营养和提高植物的抗病性来促进植物的生长, 可能与植物的糖代谢有关[28]。镰刀菌属是硒砂瓜连作土壤中主要的有害属[29], 是引起枯萎病的主要病害。此外, 研究发现青霉属微生物在不同连作年限的土壤中均有分布, 其中在T20中含量最高, 而青霉属真菌是自然界中生物有机成分的重要分解者, 能产生多种有活性的次生代谢产物, 以抵抗真菌、细菌等[30]。土壤微生物群落的组成主要是由土壤本身的性质以及耕作方式、作物类型和管理模式等因素决定[21]。近年来, 不少研究认为多年在同一块地种植相同作物, 会造成有益微生物数量减少及病原菌数量增加, 长此以往会影响植物的健康生长、正常发育等生命活动。如华菊玲等[31]研究表明连作会导致芽孢杆菌属()微生物数量减少, 而尖孢镰刀菌和青枯劳尔氏菌()数量显著高于未连作土壤。除土壤环境中微生物数量变化, 土壤理化性质的改变也会影响土壤微生物区系的变化。焦润安等[32]研究发现连作可引起土壤理化性质恶化, 土壤养分失衡, 酚酸类物质积累, 最终导致微生物群落改变, 降低土壤的健康状况, 影响马铃薯(L.)生长。在长期连作过程中, 病原菌以菌丝和分生孢子在植物残骸或土壤中越冬, 在土壤中长久生存。本研究中, T5土壤中镰刀菌属丰度最低, 假霉样真菌属丰度最高; 在T10中镰刀菌属丰度最高, 而假霉样真菌属几乎消失。由此可推测假霉样真菌属与镰刀菌属病原菌有一定相互作用, 其次与细菌群落中一些有益菌属丰度增加有关, 还需进一步验证。
3.3 土壤理化性质对真菌群落结构的影响
在连作条件下, 土壤理化性质能够更好地体现土壤的健康状况, 并且能够影响土壤微生物种群数量及分布, 不同植被的营养代谢活动导致土壤中养分具有一定的差异[3]。在本研究中, 随着连作时间延长, 土壤理化性质与连作年限间并没有显著的相关性, 但土壤理化性质与各真菌群落间有一定的相关性。Song等[6]研究黄连(Franch.)连作结果表明, 环境理化性质可直接或者间接影响真菌的生长, 从而影响真菌群落。有研究表明, 土壤理化因子与土壤微生物群落显著相关, 土壤环境因子的改变, 会影响土壤中微生物群落结构[33-34]。本研究中, 全磷、碱解氮、有效磷是影响连作硒砂瓜土壤真菌群落的主要因子, 各理化因子会显著影响相关真菌微生物群落, 其中全磷、碱解氮与镰刀菌属呈现显著的负相关, 说明全磷和碱解氮含量的增加会导致镰刀菌属丰度的减少, 因此, 推测在进行病原菌防治过程中, 可增加土壤中磷和氮含量抑制镰刀菌属病原菌的生长, 目前还没有通过改变土壤理化性质来防治病原菌的研究, 还需要进一步证明。
4 结论
随着连作时间的增加, 土壤中真菌多样性指数与丰富度指数先增加后减小, 连作改变了真菌群落在门和属水平的群落组成, 全磷、碱解氮、有效磷含量是影响土壤真菌群落最主要的因子。土壤理化性质与硒砂瓜连作年限没有显著的相关性, 与土壤中真菌群落结构存在显著的相关性, 土壤养分的变化会显著影响真菌微生物群落结构。通过研究硒砂瓜连作对土壤真菌群落结构的影响, 为了解硒砂瓜连作条件下微生群落多样性奠定基础, 也为后续研究连作障碍的生物防治提供研究基础和理论依据。
[1] TEDERSOO L, BAHRAM M, PÕLME S, et al. Global diversity and geography of soil fungi[J]. Science, 2014, 346(6213): 1256688
[2] 商鸿生. 现代植物免疫学[M]. 北京: 中国农业出版社, 2013 SHANG H S. Modern Immunology of Plants[M]. Beijing: China Agriculture Press, 2013
[3] 李锐, 刘瑜, 褚贵新, 等. 棉花连作对北疆土壤酶活性、致病菌及拮抗菌多样性的影响[J]. 中国生态农业学报, 2015, 23(4): 432–440 LI R, LIU Y, CHU G X, et al. Response of soil enzyme activity and microbial community structure, diversity to continuous cotton cropping in northern Xinjiang[J]. Chinese Journal of Eco-Agriculture, 2015, 23(4): 432–440
[4] 蒋婧, 宋明华. 植物与土壤微生物在调控生态系统养分循环中的作用[J]. 植物生态学报, 2010, 34(8): 979–988 JIANG J, SONG M H. Review of the roles of plants and soil microorganisms in regulating ecosystem nutrient cycling[J]. Chinese Journal of Plant Ecology, 2010, 34(8): 979–988
[5] 赵帆. 不同连作年限草莓根际土壤微生物多样性变化[D]. 合肥: 安徽大学, 2018 ZHAO F. Biodiversity of microorganism in rhizosphere soil of strawberry with different continuous cropping years[D]. Hefei: Anhui University, 2018
[6] SONG X H, PAN Y, LI L Y, et al. Composition and diversity of rhizosphere fungal community inFranch. continuous cropping fields[J]. PLoS One, 2018, 13(3): e0193811
[7] 纪莉景, 王连生, 栗秋生, 等. 西瓜枯萎病菌分离及致病力测定[J]. 河北农业科学, 2010, 14(8): 137–138 JI L J, WANG L S, LI Q S, et al. Isolation and pathogenicity determination of watermelon wilt[J]. Journal of Hebei Agricultural Sciences, 2010, 14(8): 137–138
[8] BOUGHALLEB N, EL MAHJOUB M. Frequency ofF. sp.and.F. sp.from watermelon seeds and their effect on disease incidence[J]. Research Journal of Parasitology, 2007, 2(1): 32–38
[9] LING N, DENG K Y, SONG Y, et al. Variation of rhizosphere bacterial community in watermelon continuous mono-cropping soil by long-term application of a novel bioorganic fertilizer[J]. Microbiological Research, 2014, 169(7/8): 570–578
[10] 袁旭东. 中宁县无公害硒砂瓜机械化生产技术探讨[J]. 当代农机, 2017, (11): 61–63 YUAN X D. Discussion on mechanized production technology of pollution-free selenium sand melon in Zhongning County[J]. Contemporary Farm Machinery, 2017, (11): 61–63
[11] 单治彬, 孔金玲, 张永庭, 等. 面向对象的特色农作物种植遥感调查方法研究[J]. 地球信息科学学报, 2018, 20(10): 1509–1519 SHAN Z B, KONG J L, ZHANG Y T, et al. Remote sensing investigation method of object-oriented crops with special characteristics[J]. Journal of Geo-Information Science, 2018, 20(10): 1509–1519
[12] 刘苹, 赵海军, 万书波, 等. 连作对花生根系分泌物化感作用的影响[J]. 中国生态农业学报, 2011, 19(3): 639–644 LIU P, ZHAO H J, WAN S B, et al. Effect of continuous cropping on allelopathy of peanut root exudates[J]. Chinese Journal of Eco-Agriculture, 2011, 19(3): 639–644
[13] BOLGER A M, LOHSE M, USADEL B. Trimmomatic: A flexible trimmer for Illumina sequence data[J]. Bioinformatics, 2014, 30(15): 2114–2120
[14] EDGAR R C. Search and clustering orders of magnitude faster than BLAST[J]. Bioinformatics, 2010, 26(19): 2460–2461
[15] EDGAR R C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads[J]. Nature Methods, 2013, 10(10): 996–998
[16] DESHPANDE V, WANG Q, GREENFIELD P, et al. Fungal identification using a Bayesian classifier and the Warcup training set of internal transcribed spacer sequences[J]. Mycologia, 2016, 108(1): 1–5
[17] KNUDSEN I M B, HOCKENHULL J, JENSEN D F. Biocontrol of seedling diseases of barley and wheat caused byand: Effects of selected fungal antagonists on growth and yield components[J]. Plant Pathology, 1995, 44(3): 467–477
[18] 王娟英, 许佳慧, 吴林坤, 等. 不同连作年限怀牛膝根际土壤理化性质及微生物多样性[J]. 生态学报, 2017, 37(17): 5621–5629 WANG J Y, XU J H, WU L K, et al. Analysis of physicochemical properties and microbial diversity in rhizosphere soil ofunder different cropping years[J]. Acta Ecologica Sinica, 2017, 37(17): 5621–5629
[19] 阎海涛, 殷全玉, 丁松爽, 等. 生物炭对褐土理化特性及真菌群落结构的影响[J]. 环境科学, 2018, 39(5): 2412–2419 YAN H T, YIN Q Y, DING S S, et al. Effect of biochar amendment on physicochemical properties and fungal community structures of cinnamon soil[J]. Environmental Science, 2018, 39(5): 2412–2419
[20] 赵帆, 赵密珍, 王钰, 等. 不同连作年限草莓根际细菌和真菌多样性变化[J]. 微生物学通报, 2017, 44(6): 1377–1386 ZHAO F, ZHAO M Z, WANG Y, et al. Biodiversity of bacteria and fungi in rhizosphere of strawberry with different continuous cropping years[J]. Microbiology China, 2017, 44(6): 1377–1386
[21] MAZZOLA M, MANICI L M. Apple replant disease: Role of microbial ecology in cause and control[J]. Annual Review of Phytopathology, 2012, 50: 45–65
[22] 康亚龙, 孙文庆, 刘建国, 等. PLFA方法研究连作对加工番茄根际土壤微生物群落结构的影响[J]. 中国生态农业学报, 2017, 25(4): 594–604KANG Y L, SUN W Q, LIU J G, et al. Microbial community structure in rhizosphere soils of long-term continuously cropped processing tomato based on PLFA method[J]. Chinese Journal of Eco-Agriculture, 2017, 25(4): 594–604
[23] SCHOCH C L, SUNG G H, LÓPEZ-GIRÁLDEZ F, et al. The Ascomycota tree of life: A phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits[J]. Systematic Biology, 2009, 58(2): 224–239
[24] Burnham-Marusich A R, Breeana H, Kvam A J, et al. Conservation of Mannan Synthesis in fungi of the Zygomycota and Ascomycota reveals a broad diagnostic target[J]. mSphere, 2018, 3(3): e00094-18
[25] OLDROYD G E D. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants[J]. Nature Reviews Microbiology, 2013, 11(4): 252–263
[26] GOMES N C M, FAGBOLA O, COSTA R, et al. Dynamics of fungal communities in bulk and maize rhizosphere soil in the tropics[J]. Applied and Environmental Microbiology, 2003, 69(7): 3758–3766
[27] ZHANG H S, WU X H, LI G, et al. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (sp.) and their effects ongrowth and enzyme activities of rhizosphere and bulk soils at different salinities[J]. Biology and Fertility of Soils, 2011, 47(5): 543–554
[28] LIAO X G, FANG W G, LIN L C, et al.produces an extracellular invertase (MrINV) that plays a pivotal role in rhizospheric interactions and root colonization[J]. PLoS One, 2013, 8(10): e78118
[29] MANICI L M, KELDERER M, FRANKE-WHITTLE I H, et al. Relationship between root-endophytic microbial communities and replant disease in specialized apple growing areas in Europe[J]. Applied Soil Ecology, 2013, 72: 207–214
[30] 罗清, 彭程, 叶波平. 青霉属真菌研究新进展[J]. 药物生物技术, 2016, 23(5): 452–456 LUO Q, PENG C, YE B P. New advances in research of the genus[J]. Pharmaceutical Biotechnology, 2016, 23(5): 452–456
[31] 华菊玲, 刘光荣, 黄劲松. 连作对芝麻根际土壤微生物群落的影响[J]. 生态学报, 2012, 32(9): 2936–2942 HUA J L, LIU G R, HUANG J S. Effect of continuous cropping of sesame on rhizospheric microbial communities[J]. Acta Ecologica Sinica, 2012, 32(9): 2936–2942
[32] 焦润安, 徐雪风, 杨宏伟, 等. 连作对马铃薯生长和土壤健康的影响及机制研究[J]. 干旱区农业研究, 2018, 36(4): 94–100 JIAO R A, XU X F, YANG H W, et al. Study on the effect and its mechanism of continuous cropping on potato growth and soil health[J]. Agricultural Research in the Arid Areas, 2018, 36(4): 94–100
[33] 秦越, 马琨, 刘萍. 马铃薯连作栽培对土壤微生物多样性的影响[J]. 中国生态农业学报, 2015, 23(2): 225–232 QIN Y, MA K, LIU P. Effect of potato continuous cropping on genetic diversity of soil microorganisms[J]. Chinese Journal of Eco-Agriculture, 2015, 23(2): 225–232
[34] HUHE, CHEN X J, HOU F J, et al. Bacterial and fungal community structures in loess plateau grasslands with different grazing intensities[J]. Frontiers in Microbiology, 2017, 8: 606
The effect of continuous cropping of selenium melon on soil fungal community structure*
FENG Cui’e, YUE Sijun**, JIAN Ani, CHEN Liping, GUO Yang, ZHENG Rui, SU Jianyu**
(College of Life Sciences, Ningxia University / Key Laboratory of Ministry of Education for Conservation and Utilization of Special Biological Resources in the Western China, Yinchuan 750021, China)
Selenium-melon is an important cash crop in Ningxia, but its yield and quality decrease when continuously cropped. The decrease may be related to soil fungal community changes that take place during continuous cropping, which was determined in this study. Soil samples from continuous cropping of selenium-melon were analyzed using Illumina Miseq high-throughput sequencing. The analysis showed that, after an initial increase, the diversity and density of the fungal community in the soil of continuous cropping decreased. Eight fungal phyla, 25 classes, and 244 genera were identified in the soil samples. Ascomycota and Zygomycota were the predominant phyla, accounting for > 90% of the fungi. The abundance decreased 32.51% for Ascomycota, but increased 29.89% for Zygomycota, after 30 years of continuous cropping. The most predominant genera were,,,and, and the abundance changes included a 45.81% increase forafter 5 years, a 26.74% increase forafter 10 years, a 26.83% decrease forafter 15 years, a 29.68% increase forafter 20 years, a 18.30% decrease forafter 25 years, and a 29.89% increase forafter 30 years of continuous cropping. Soil physical and chemical properties were not related to years of continuous cropping, but affected the fungal community structure. Soil total phosphorus, alkaline nitrogen, and available phosphorus were the most influential factors for fungal genus changes. A significant positive correlation was observed between the abundance ofand soil total potassium, between the abundance ofand soil total phosphorus and available potassium, and between the abundance ofand soil pH, whereas a significant negative correlation was observed between the abundance ofand soil total phosphorus and alkali-hydrolyzed nitrogen. Diminished production of selenium-melon in continuous cropping was related to soil fungal community changes rather than soil physicochemical properties, demonstrating that selenium-melon soil-borne diseases might be controlled through biological means.
Continuous cropping; Soil fungi; Community structure; High-throughput sequencing; Soil physicochemical properties; Selenium melon
YUE Sijun, E-mail: sijunyue@126.com; SU Jianyu, E-mail: su_jy@nxu.edu.cn
Aug. 7, 2018;
Nov. 26, 2018
10.13930/j.cnki.cjea.180746
S154.3
A
2096-6237(2019)04-0537-08
岳思君, 主要研究方向为微生物学, E-mail: sijunyue@126.com; 苏建宇, 主要研究方向为工业微生物, E-mail: su_jy@nxu.edu.cn
冯翠娥, 主要研究方向为微生物技术与工程。E-mail: 13519291697@163.com
2018-08-07
2018-11-26
* This study was supported by the National Natural Science Foundation of China (31360025, 31560418) and Ningxia Key Research and Development Program (2017BY081).
* 国家自然科学基金项目(31360025, 31560418)和宁夏回族自治区重点研发计划项目(2017BY081)资助
冯翠娥, 岳思君, 简阿妮, 陈丽萍, 郭洋, 郑蕊, 苏建宇. 硒砂瓜连作对土壤真菌群落结构的影响[J]. 中国生态农业学报(中英文), 2019, 27(4): 537-544
FENG C E, YUE S J, JIAN A N, CHEN L P, GUO Y, ZHENG R, SU J Y. The effect of continuous cropping of selenium melon on soil fungal community structure[J]. Chinese Journal of Eco-Agriculture, 2019, 27(4): 537-544
