短期生物炭添加对不同类型土壤细菌和氨氧化微生物的影响
2019-04-12张梦阳夏浩吕波丛铭宋文群姜存仓
张梦阳,夏浩,吕波,丛铭,宋文群,姜存仓
短期生物炭添加对不同类型土壤细菌和氨氧化微生物的影响
张梦阳1,夏浩1,吕波1,丛铭1,宋文群2,姜存仓1
(1华中农业大学资源与环境学院微量元素研究中心,武汉 430070;2江西省信丰县农业技术推广站,江西赣州 341600)
【目的】氨氧化作用是硝化作用的第一步,也是硝化作用的限速步骤,是全球氮循环的关键环节。本试验旨在研究在我国不同类型土壤中添加花生壳生物炭对细菌和氨氧化作用的影响,为生物炭的推广使用提供理论依据。【方法】试验以黄棕壤、潮土、黑土为供试土壤,通过短期培养试验,利用16SrRNA测序研究生物炭对不同类型土壤氨氧化微生物、细菌群落结构以及相关酶基因表达量的影响。每种土壤设置4个处理:CK(不施用化肥和生物炭),F(单施化肥),C(单施2%花生壳生物炭),FC(施用化肥+2%花生壳生物炭)。【结果】施用生物炭后(C、FC)酸性土壤pH显著提高了0.5—1.0个单位,但碱性土壤pH显著降低了0.5—0.6个单位;单施生物炭(C)造成黄棕壤的微生物丰富度和多样性显著提高,潮土在单施生物炭(C)时仅显著提高了土壤的微生物多样性指数,在黑土中施用生物炭和化肥都未显著改变土壤微生物的丰富度和多样性;在3种土壤中氨氧化细菌的丰度皆高于氨氧化古菌,测得的氨氧化细菌的OTU丰度约为氨氧化古菌的8.1倍;生物炭和化肥并未显著改变奇古菌门中的OTU丰度,却对β和γ变形菌中的OTU丰度产生了显著性影响;3种土壤的氨氧化细菌都以β变形菌为主,约占60%;另外,生物炭的施用(C、FC)在PC1(40.4%)上显著改变了黄棕壤的微生物群落结构,在PC1(42.3%)和PC2(21.3%)上都显著改变了潮土的微生物群落结构;施用生物炭后(C、FC),短期内潮土中氨合成相关酶基因表达量显著降低14.7%—39.9%,氨氧化古菌丰度在单一施炭(C)和化肥与生物炭同施(FC)时分别降低了70.5%和48.7%。【结论】施用生物炭后,短期内显著改变了黄棕壤和潮土的微生物群落结构,并明显抑制了潮土的氨氧化作用。
生物炭;化肥;氨氧化作用;微生物群落结构;黄棕壤;潮土;黑土
0 引言
【研究意义】氮是组成生命最重要物质核酸和蛋白质必不可少的元素之一[1],土壤圈是氮循环中最活跃的区域,土壤中氮循环对土壤质量、农业生产力乃至其他地球化学循环都有着重要的作用,而微生物是必不可少的推动者[2-3]。生物炭近年来被广泛应用于改良土壤、提高土壤固炭能力等[4],但生物炭施用于不同土壤时改良效果不同[5],生物炭在土壤中的具体微生物效应也不明确。因此,研究生物炭对不同类型土壤微生物群落和氨氧化作用在农业生产中意义重大。【前人研究进展】生物炭作为一种土壤改良剂,郑瑞伦等[6]的研究表明,400℃热解得到的秸秆炭施用于壤土中显著改善土壤pH、通气性、含水量、有效养分等。李昌见等[7]通过大田试验也发现施用10 t·hm-2到60 t·hm-2玉米秸秆炭都显著提高了砂壤土的含水量、肥料利用率。吕波等[8]施加3%生物炭后发现红壤和黄棕壤的pH、速效养分以及酶活性都得到显著提高。应介官等[9]研究也表明在红壤中添加生物炭可以显著降低土壤铝毒。相应研究也证明了在不同温度下热解得到的生物炭都不同程度降低营养物质的淋失,ASADA等[10-11]研究表明,低温(400—500℃)热解产生的毛竹生物炭有更强的NH3吸附能力。CLOUGH等[12]发现,高于600℃热解产生的木材生物炭才具有较好的NO3-吸附能力。前人研究也发现生物炭的施用也与温室气体的排放有关,有学者利用9种不同原料经500℃热解得到的生物炭,研究了它们在14种不同类型土壤中N2O的排放,结果表明施用生物炭提高了土壤pH,从而减少土壤中N2O的排放[13],但也有研究发现施用600℃热解得到的木质生物炭对草地土壤N2O排放无影响[12],甚至有研究者发现无论是高温(900℃)生产的核桃炭还是中温(550℃)生产松木炭都促进砂质黏壤土N2O的排放[14]。氨氧化作用为硝化作用的限速步骤,该作用主要由氨氧化微生物推动,氨氧化细菌广泛的分布在土壤生态系统、淡水生态系统和海洋生态系统中,且氨氧化细菌主要属于β变形菌纲和γ变形菌纲[15-16]。氨氧化古菌的发现让我们了解到参与氨氧化作用的不仅有细菌还有古菌。最初,人们认为氨氧化古菌隶属于温泉古菌(Crenarchaeota)[17],BrochierArmanet等[18]研究并提出了新的古菌菌门奇古菌门(Thaumarchaeota)。也有研究证明奇古菌几乎包括了所有发现的土壤、海洋、热泉中的氨氧化古菌[19]。目前来看,生物炭对土壤氮循环的影响已开展较多研究,并发现生物炭的添加可能影响着氮素淋溶、氨挥发、硝化作用、固氮作用、以及氧化亚氮的产生[20-24],微生物驱动土壤氮循环的各过程[2-3]。我们可以明确土壤类型差异是导致生物炭施用效果差异产生的主要原因之一。ABUJABHAH等[25]也发现热解温度为650—750℃的桉树生物炭对土壤微生物群落和氮循环细菌的影响主要取决于土壤类型,其中对红壤和黑土的作用明显高于棕壤。LIU等[26]施用了0—40 t·hm-2的稻壳生物炭(400℃)于红壤、棕壤和盐渍土中,结果发现生物炭对红壤细菌群落改变最显著。【本研究切入点】在不同土壤中驱动土壤氨氧化作用的主要微生物可能是氨氧化细菌也可能是氨氧化古菌,但生物炭的添加究竟是改变了土壤中氨氧化细菌的丰度还是氨氧化古菌的丰度;在不同土壤施用生物炭后氨氧化微生物的变化趋势是否相同;生物炭对酸碱性不同的土壤造成的氨氧化微生物的具体差异是否相同,很多类似的问题仍没有明确的答案。【拟解决的关键问题】本文选取我国3种不同类型土壤(黄棕壤、潮土、黑土)为供试土壤,通过16S rRNA测序分析得到单一施炭、单一施肥、生物炭与化肥同施对不同类型土壤细菌群落结构、氨氧化微生物丰度以及氮代谢通路中相关酶基因表达量的具体差异,从而揭示生物炭对不同土壤细菌群落和氨氧化作用的具体影响,为生物炭的实际应用提供理论依据。
1 材料与方法
1.1 样本采集及试验处理
试验土壤采集湖北黄棕壤、山东潮土、黑龙江黑土,每个地区选择100 m2的区域,在区域中随机选择10个采样点采集0—20 cm土层的土壤样品,分别代表酸性、弱酸性和碱性土壤(供试土壤基本性质见表1)。样品自然风干后挑去石子、植物根系等杂物,全部研磨过2 mm筛,混匀后储存备用。设4个处理:CK(不施用化肥和生物炭),F(单施化肥),C(单施生物炭),FC(施用化肥+生物炭)。化学肥料施用KH2PO4,KNO3,NH4NO3和Ca(NO3)2(其中N 0.46 g·kg-1干土,P2O50.07 g·kg-1干土,K2O 0.24 g·kg-1干土)。沈阳农业大学提供花生壳生物炭,生物炭在400℃下热解制备而成,全炭321.9 g·kg-1,全氮18.8 g·kg-1,全磷2.6 g·kg-1,全钾8.5 g·kg-1,生物炭施用量为2%(生物炭质量/土壤质量)。试验在塑料杯中进行培养,每杯装土300 g,根据试验方案加入化肥和生物炭并混匀,土壤上层平铺一层滤纸防止浇水时扰动土壤,每个处理设置3次重复,试验过程中土壤持水量保持在40%左右。土壤与肥料和生物炭按处理混匀后加水至饱和持水量,静置熟化1周,熟化后培养4周,试验于2018年5月6日至6月1日在华中农业大学盆栽场进行。试验结束后采集新鲜土壤于-80℃保存用于DNA的提取以及测定,其余土壤自然风干后,过1 mm筛,用于测定土壤pH(液土比例为2.5﹕1)。

表1 供试土壤的基本性质
1.2 DNA 抽提和PCR扩增
根据E.Z.N.A.® soil试剂盒(Omega Bio-tek,Norcross,GA,U.S.)说明书进行总DNA抽提,DNA浓度和纯度利用NanoDrop2000进行检测,利用1%琼脂糖凝胶电泳检测DNA提取质量;用515F(5′-GT GCCAGCMGCCGCGG-3′)和806R(5′-GGACTACH VGGGTWTCTAAT-3′)引物对V4可变区进行PCR扩增。
1.3 Illumina Miseq测序
使用2%琼脂糖凝胶回收PCR产物,利用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,Union City,CA,USA)进行纯化,Tris-HCl洗脱,2%琼脂糖电泳检测。利用QuantiFluor™-ST(Promega,USA)进行检测定量。根据Illumina MiSeq 平台(Illumina,San Diego,USA)标准操作规程将纯化后的扩增片段构建PE 2*300的文库。
1.4 数据处理
原始测序序列使用Trimmomatic 软件质控,使用FLASH软件进行拼接[27]:
(1)设置50 bp的窗口,如果窗口内的平均质量值低于20,从窗口前端位置截去该碱基后端所有序列,之后再去除质控后长度低于50 bp的序列;
(2)根据重叠碱基overlap将两端序列进行拼接,拼接时overlap之间的最大错配率为0.2,长度需大于10 bp。去除无法拼接的序列。
(3)根据序列首尾两端的barcode和引物将序列拆分至每个样本,barcode需精确匹配,引物允许2个碱基的错配,去除存在模糊碱基的序列。
本研究使用UPARSE软件(version 7.1 http:// drive5.com/uparse/),根据97%的相似度对序列进行OTU聚类,并在聚类的过程中去除单序列和嵌合体[28]。利用RDP classifier (http://rdp.cme.msu.edu/)对每条序列进行物种分类注释,比对Silv128/16S数据库,设置比对阈值为70%[29]。分析所用序列为原始序列按照最小样本序列数进行抽平后得到。文章中物种组成柱状图基于tax_summary_a文件夹中的数据表,利用R语言工具作图,用于分析处理间物种组成差异,物种丰度<0.005合并为Other;Alpha多样性指数利用mothur(version v.1.30.1 http://www.mothur.org/ wiki/ Schloss_SOP#Alpha_diversity)指数分析[30],指数组间差异检验方法采用Student’s T检验。β多样性利用R语言PCA统计分析和作图。16S功能预测通过PICRUSt去除16S marker gene在物种基因组中的copy数目的影响,通过比对KEGG获得代谢通路中的各个层级的代谢信息以及相关酶的基因表达量[31]。
采用Microsoft Excel 2010对数据整理,用SPSS 20.0软件进行单因素方差和多因素分析,ANOVA进行显著性差异比较,多重比较采用Duncan检验,所有数值均为3次重复的平均值。
2 结果
2.1 生物炭对不同土壤pH的影响
图1所示,在黄棕壤中施用生物炭后显著提升了其土壤的pH,提升了0.6—1.0个单位,然而单一施用化肥并未产生显著性差异。在潮土中施用生物炭pH提升了0.5—0.6个单位,而仅施用化肥时降低了0.3个单位。在黑土中,与CK相比,无论是施用生物炭还是化肥都造成了土壤pH的显著降低,降幅为0.2—0.6个单位。通过三因素方差分析发现,无论单一施肥、单一施炭还是生物炭与化肥同施都对土壤pH产生显著影响,另外三因素两两间的交互效应也对土壤pH存在显著影响,但三因素间的交互效应并无显著影响(表2)。
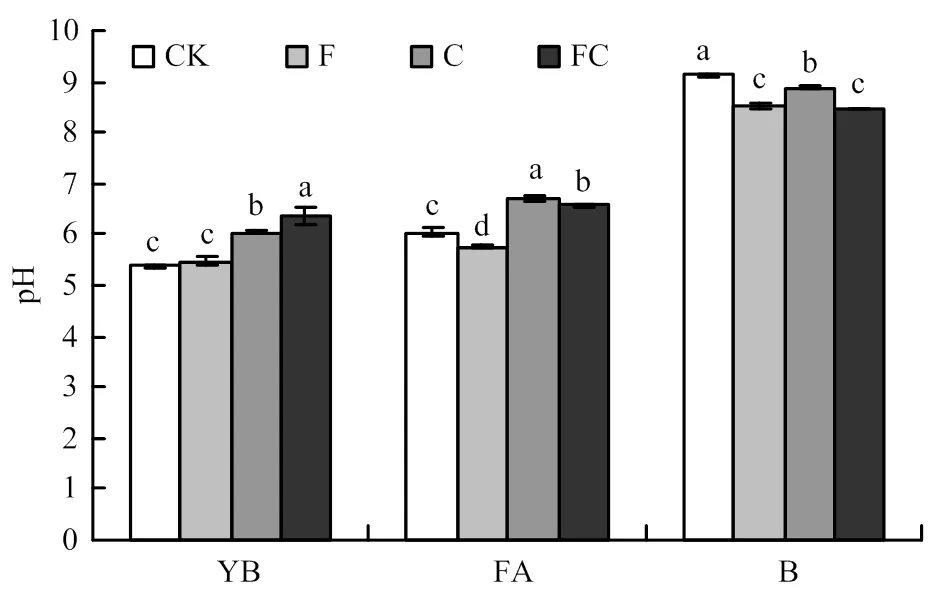
YB:黄棕壤;FA:潮土;B:黑土。CK,F,C,FC分别代表对照,单施化肥,单施生物炭和生物炭与化肥同施4种处理。图柱上不同字母表示在处理间存在显著差异(P<0.05)。下同

表2 土壤pH三因素方差分析
2.2 生物炭对不同土壤细菌、古菌丰富度和多样性指数的影响
本研究采用Ace指数和shannon指数表示微生物丰富度指数以及多样性指数,与对照相比分析经化肥或生物炭处理后的指数变化(图2)。在黄棕壤中单施生物炭提升了土壤细菌和古菌的丰富度和多样性,且与对照、单施化肥处理之间存在显著性差异,单施化肥降低了黄棕壤的丰富度和均匀度指数,但无显著性差异,经炭肥同施后也并未出现显著变化(图2-a,图2-b)。在潮土中,单施化肥降低了土壤细菌和古菌的丰富度和多样性,但仅在丰富度指数上存在显著性差异,单施生物炭后细菌和古菌的多样性指数得到显著提升,生物炭与化肥同施后细菌和真菌的丰富度和多样性指数变化幅度均较小(图2-c,2-d)。在黑土中,不同处理对土壤细菌和古菌丰富度和多样性指数未出现显著性改变(图2-e,2-f)。
2.3 生物炭对不同土壤氨氧化微生物丰度的影响
通过分析测序所得氨氧化微生物的OTU丰度发现,无论是在黄棕壤还是在潮土和黑土中,氨氧化细菌的OTU丰度远大于氨氧化古菌的OTU丰度。另外,氨氧化古菌经处理后测得的OTU丰度差异性不显著,而氨氧化细菌在处理后则出现了显著性差异(表3)。
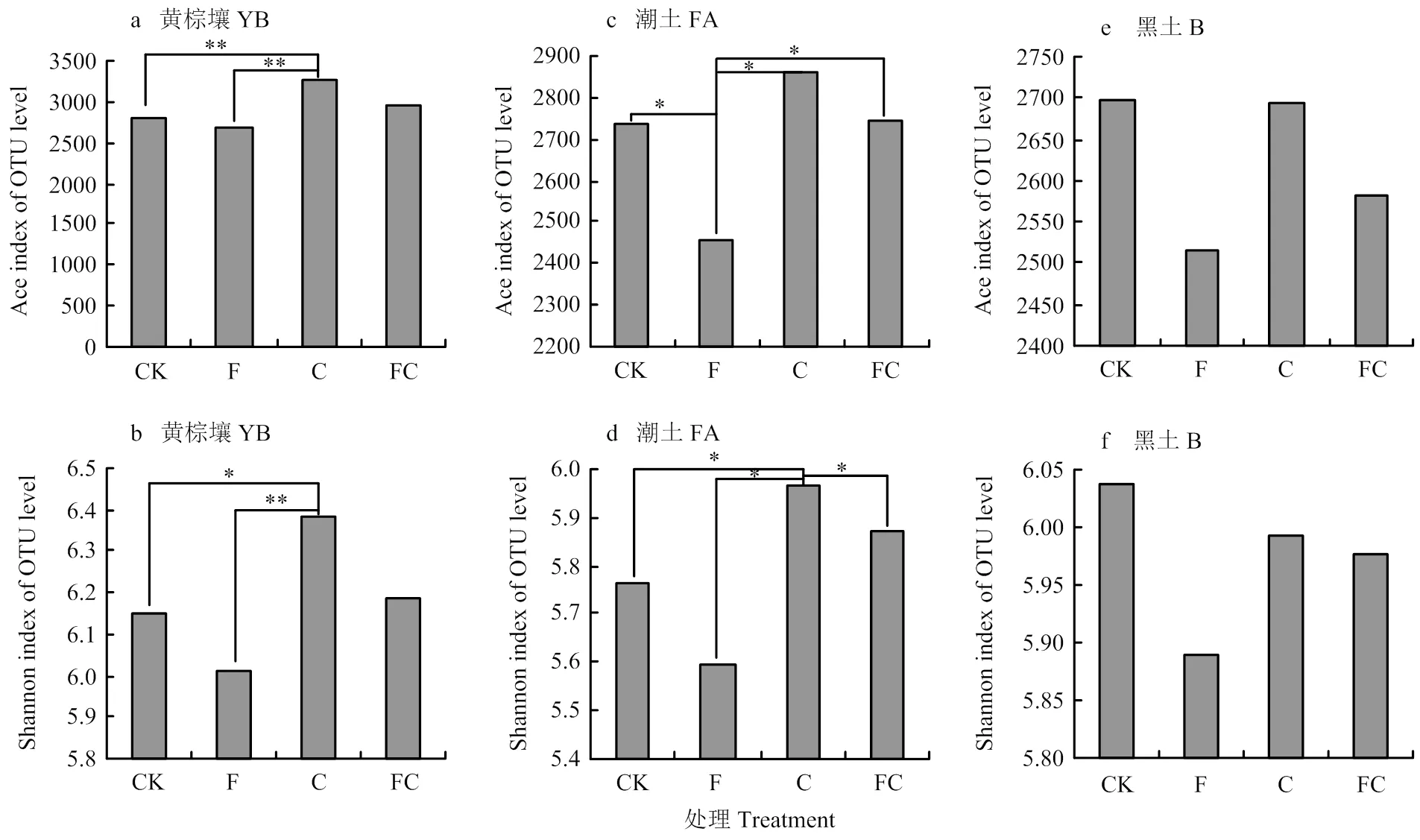
*代表在0.05水平下差异显著,**代表在0.01水平下差异显著

表3 氨氧化微生物OTU丰度
为进一步了解土壤中氨氧化微生物丰度的变化趋势,针对古菌中的奇古菌和细菌中的β变形菌和γ变形菌的丰度变化进行了分析(图3)。结果显示,在黄棕壤氨氧化古菌丰度变化上,奇古菌门在单施化肥和单施生物炭后分别提升了约81.8%和45.6%,但在生物炭与化肥同施时,其丰度下降了约15.0%(图3-a)。在氨氧化细菌丰度变化上,主要以β变形菌为主,约占92.3%。β和γ变形菌之和以生物炭与化肥同施处理最高,与对照相比提高了14.8%,而单施生物炭却降低了18.0%(图3-b)。在潮土中,所有处理对土壤奇古菌门的丰度均存在降低作用,其中施炭处理(F和FC)的降低趋势最明显,分别降低了70.5%和48.7%(图3-c)。另外,潮土中氨氧化细菌主要以β变形菌为主(图3-d)。与潮土不同的是,黑土中所有处理对奇古菌门的丰度都存在提升趋势,其中施炭处理(F和FC)提升最明显,分别提高了66.7%和48.1%(图3-e)。在氨氧化细菌方面,虽然黑土也以β变形菌为主,但是γ变形菌的丰度比例远高于其余潮土和黄棕壤。另外,在炭肥同施时黑土中氨氧化细菌的丰度也出现较大幅度的升高(图3-f)。
2.4 生物炭对不同土壤细菌、古菌群落结构的作用
通过主成分分析细菌和古菌的β多样性,进一步探究各处理间群落结构的差异(图4)。在黄棕壤中,单施化肥样本点与对照样本点分离程度较低,表明单施化肥对土壤的细菌和古菌的群落结构影响较小。施炭处理(C和FC)与对照在X轴(40.4%)上存在明显分离,另外单施炭和炭肥同施在Y轴(19.5%)上也存在明显的分离(图4-a)。在潮土中,施炭处理(C和FC)样本点皆分布在第四象限,与对照处理在X轴(42.3%)和Y轴(21.3%)上都存在明显分离(图4-b)。在黑土中,对照、单施化肥以及炭肥同施三者之间没有出现明显的分离,但是单施化肥样本点与对照样本点间距离相对较远,说明在黑土中单施化肥对土壤细菌和真菌群落结构的影响相对较大(图4-c)。

图3 不同土壤各处理间奇古菌,β变形菌和γ变形菌的丰度变化柱状图

图4 不同土壤各处理间主成分分析
2.5 生物炭对土壤氨合成相关酶和氨氧化作用相关酶基因表达量的影响
土壤中氮素循环过程中需要通过各个支路中各种各样的酶相互协调完成(图5,数据网址:http://www. genome.jp/kegg/pathway.html)。根据氮素代谢通路丰度表可以发现,黄棕壤和黑土经化肥或生物炭处理后氮素代谢通路丰度并未发生显著性改变,但在潮土中添加生物炭后氮素代谢通路丰度出现显著性降低(表4)。

图5 KEGG氮素代谢通路
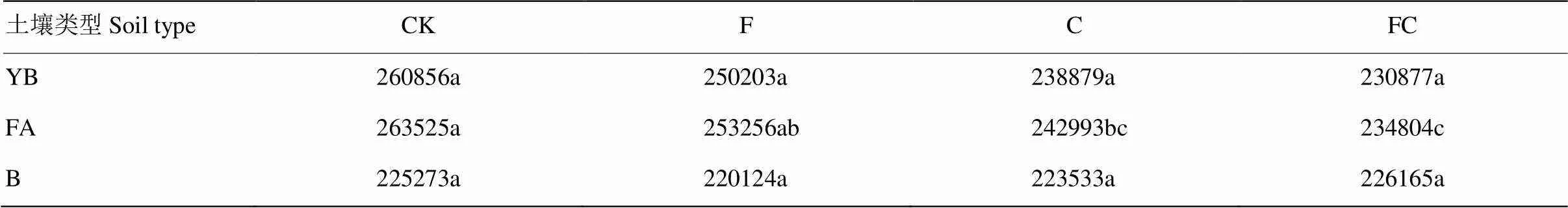
表4 不同土壤各处理的氮代谢通路丰度
根据氮素代谢通路图可以得到与氮素代谢相关酶和基因的对应关系,并通过比对数据库得到一些与氨合成和氨氧化作用相关酶的相关表达(表5)。分析氮素代谢通路中相关酶基因表达量,研究在不同土壤不同处理中相关酶的变化情况,发现检测到的酶在黑土中出现变化的较少,只有甲酰胺酶(1.7.2.1)和腈水解酶(3.5.1.49)在处理间存在显著差异。然而在黄棕壤和潮土中存在显著差异的酶数量明显多于黑土,且不同土壤间经化肥或生物炭处理后出现显著性差异的酶的种类也存在明显区别。
黄棕壤中,经化肥或生物炭处理后,与对照相比,氨合成和氨氧化作用相关酶的基因表达量有不同程度的改变,但均未达到显著差异。在潮土中,生物炭的施用(F、FC)显著降低了谷氨酸脱氢酶(2.7.2.2),谷氨酸脱氢酶(NAD(P)+)(1.4.1.2),谷氨酸脱氢酶(NADP+)(1.4.1.3)的表达量,另外,单施化肥也显著降低了谷氨酸脱氢酶(NADP+)(1.4.1.3)的表达量,但会显著提升亚硝酸还原酶(不分解)(1.4.1.4)的基因表达量。在黑土中经不同处理后基因表达量出现显著性变化的酶种类较少,甲酰胺酶(1.7.2.1)在炭肥同施时表达量显著提升,而氰酸裂合酶(3.5.5.1)在单施生物炭时表达量显著下降。

表5 土壤氨合成相关酶和氨氧化作用相关酶基因表达量
3 讨论
在本研究中酸性土壤施用生物炭后pH显著提高了0.6—1.0个单位,但酸性土壤的氨氧化作用不仅未发现显著提高,还出现了下降的趋势,黄棕壤氮代谢通路丰度约降低了8.4%,而潮土中奇古菌的丰度约降低了70.5%,氮代谢通路丰度也显著降低了7.8%,其中在潮土中呈现显著下降的趋势,这或许是因为本试验培养时间较短。另外,土壤pH是影响氨氧化过程相关氨氧化微生物组成及群落结构的重要因素[32],在本研究中,这3种土壤中测得β变形菌和γ变形菌数量远高于奇古菌数量,表明细菌在这三种土壤的氨氧化作用中占主导地位。很多研究都表明大多数酸性土壤中,氨氧化古菌的数量要高于氨氧化细菌,因为氨氧化古菌对低pH有更强的适应能力[33-36]。本试验采用的3种土壤分别为酸性,弱酸性以及碱性土壤,单施化肥对土壤pH的影响比生物炭弱,生物炭的添加显著提升了酸性土壤的pH,但是土壤依旧呈酸性,而黑土在施用生物炭后显著降低了pH,但土壤也仍呈碱性,在本研究中并未发现酸性土壤中奇古菌占优势。这可能与515F和806R作为测序引物有关,该引物可以用于鉴定细菌和古菌,但是更偏向于细菌鉴定[37],或许在古菌的鉴定上存在一定偏差。
ABUJABHAH等[25]认为热解温度为650—750℃的桉树生物炭对土壤微生物群落和氮循环细菌的影响主要取决于土壤类型,其中对施用于红壤时作用明显高于棕壤。LIU等[26]施用了0—40 t·hm-2的稻壳生物炭(400℃)于红壤、棕壤和盐渍土中,结果发现生物炭对红壤细菌群落改变最显著。通过我们的研究也发现,花生壳生物炭对黄棕壤和潮土的细菌群落结构影响更明显,这表明生物炭对偏酸性土壤微生物群落结构存在更显著的作用,主要原因是生物炭显著提高了酸性土壤的pH,从而导致细菌群落发生显著改变。很多生物炭研究围绕不同土壤氨氧化微生物的改变得到了不同的结果。HE等[38]发现热解温度为500℃的稻壳生物炭不会对碱性土壤产生显著影响,但会改变酸性土壤的氨氧化微生物丰度。李双双等[39]在黄棕壤中施用热解温度为500℃的小麦秸秆炭发现单独施用生物炭显著降低了氨氧化细菌的丰度。也有相应研究发现在复垦土壤中施用400—500℃热解成的小麦秸秆生物炭会提高土壤氨氧化细菌和古菌的丰度,并且氨氧化细菌的丰度还会随着生物炭用量的增加而增加[40]。我们的研究结果与之不同,我们发现生物炭的施用主要降低酸性土壤中氨氧化细菌的丰度,研究结果间的差异主要与土壤类型有关。BI等[41]在砂壤土中仅施用在600℃下热解得到的稻草炭后发现并未显著影响土壤氨氧化微生物,WU等[42]也在砂壤土上进行了玉米芯生物炭的试验,结果也发现仅施用生物炭并未显著改变氨氧化微生物的组成。结合前人的研究和本试验的结果可以发现,生物炭对土壤微生物的影响,因炭而异,因土而异,在使用生物炭的时候应考虑生物炭类型和土壤类型。同时也有研究者发现长期添加生物炭和短期施用生物炭的土壤中微生物群落存在明显差异[43-44]。本研究周期相对较短,与长期试验结果是否一致有待进一步探讨。
有研究认为土壤微生物群落对化肥的投入以及土壤营养条件的改变十分敏感[45],但在不同报道中的结果差异很大且没有清晰的趋势,有研究表明肥料的施用与否对细菌群落无显著影响[46-47],但也有研究表明施肥导致了微生物群落组成的改变[48-49]。本研究中,单施化肥时并未发现黄棕壤、潮土、黑土出现显著性差异。这与GEISSELER等[50]的研究结果相似,他发现绝大部分研究中土壤主要的微生物群落并没有因为化肥的施用而产生明显的改变。同样在Ultuna 和Broadbalk 的冬小麦试验中,化肥与不施肥处理相比较,细菌的群落组成并没有显著的差异[46-47,51]。
4 结论
在土壤氨氧化作用中,黄棕壤、潮土和黑土都表现为氨氧化细菌占主导地位。生物炭的添加在短期内会对黄棕壤及潮土的细菌和古菌群落结构产生显著影响。施用生物炭后,在短期内潮土的合成氨相关酶基因表达量显著降低,氨氧化古菌丰度也明显降低,从而抑制了氨氧化作用;化肥和生物炭对黄棕壤和黑土的氨氧化作用也存在一定程度的影响。
[1] CANFIELD D E, GLAZER A N, FALKOWSKI P G. The evolution and future of Earth's nitrogen cycle., 2010, 330(6001): 192-196.
[2] 李霞. 微生物在氮循环中的作用. 松辽学刊(自然科学版), 1998, 10(4): 30-33.
LI X. The role of microorganisms in the nitrogen cycle., 1998, 10(4): 30-33. (in Chinese)
[3] 贺纪正, 张丽梅. 氨氧化微生物生态学与氮循环研究进展. 生态学报, 2009, 29(1): 406-415.
HE J Z, ZHANG L M. Advances in ammonia oxidation microbial ecology and nitrogen cycle., 2009, 29(1): 406-415. (in Chinese)
[4] 陈温福, 张伟明, 孟军, 徐正进. 生物炭应用技术研究. 中国工程科学, 2011, 13(2):83-89.
CHEN W F, ZHANG W M, MENG J, XU Z J. Biochar application technology research.2011, 13(2): 83-89. (in Chinese)
[5] 陈心想, 何绪生, 耿增超, 张雯, 高海英. 生物炭对不同土壤化学性质、小麦和糜子产量的影响. 生态学报, 2013, 33(20): 6534-6542.
CHEN X X, HE X S, GENG Z C, ZHANG W, GAO H Y. Effects of biochar on different soil chemical properties, wheat and hazelnut yield.2013, 33(20): 6534-6542. (in Chinese)
[6] 郑瑞伦, 王宁宁, 孙国新, 谢祖彬, 庞卓, 王庆海, 武菊英. 生物炭对京郊沙化地土壤性质和苜蓿生长、养分吸收的影响. 农业环境科学学报, 2015, 34(5): 904-912.
ZHENG R L, WANG N N, SUN G X, XIE Z B, PANG Z, WANG Q H, WU J Y. Effects of biochar on soil properties, alfalfa growth and nutrient uptake in desertification areas of Beijing suburbs., 2015, 34(5): 904-912. (in Chinese)
[7] 李昌见, 屈忠义, 勾芒芒, 苏永莉, 霍星. 生物炭对土壤水肥利用效率与番茄生长影响研究. 农业环境科学学报, 2014, 33(11): 2187-2193.
LI C J, QU Z Y, GOU M M, SU Y L, HUO X. Effects of biochar on soil water and fertilizer utilization efficiency and tomato growth., 2014, 33(11): 2187-2193. (in Chinese)
[8] 吕波, 王宇函, 夏浩, 姚子涵, 姜存仓. 不同改良剂对黄棕壤和红壤上白菜生长及土壤肥力影响的差异. 中国农业科学, 2018, 51(22): 4306-4315.
LV B, WANG Y H, XIA H, YAO Z H, JIANG C C. Effects of Biochar and Other Amendments on the Cabbage Growth and Soil Fertility in Yellow-Brown Soil., 2018, 51(22): 4306-4315. (in Chinese)
[9] 应介官, 林庆毅, 张梦阳, 黄毅, 彭抒昂, 姜存仓. 生物炭对铝富集酸性土壤的毒性缓解效应及潜在机制. 中国农业科学, 2016, 49(23): 4576-4583.
YING J G, LIN Q Y, ZHANG M Y, HUANG Y, PENG S A, JIANG C C. Mitigative effect of biochar on aluminum toxicity of acid soil and the potential mechanism., 2016, 49(23): 4576-4583. (in Chinese)
[10] ASADA T, ISHIHARA S, YAMANE T, TOBA A, YAMADA A. Science of bamboo charcoal: Study of carbonizing temperature of bamboo charcoal and removal capability of harmful gases., 2002, 48(6): 473-479.
[11] ASADA T, OHKUBO T, KAWATA K, OIKAWA K. Ammonia adsorption on bamboo charcoal with acid treatment., 2006, 52(5): 585-589.
[12] CLOUGH T J, BERTRAM J E, RAY J L, CONDRON L M, O'CALLAGHAN M, SHERLOCK R R, WELLS N S. Unweathered wood biochar impact on nitrous oxide emissions from a bovine-urine- amended pasture soil., 2010, 74(3): 852-860.
[13] CAYUELA M L, SÁNCHEZ-MONEDERO M A, ROIG A, HANLEY K, ENDERS A, LEHMANN J. Biochar and denitrification in soils: when, how much and why does biochar reduce N2O emissions?, 2013, 3: 1732.
[14] VERHOEVEN E, SIX J. Biochar does not mitigate field-scale N2O emissions in a Northern California vineyard: an assessment across twoyears., 2014, 191(15): 27-38.
[15] PURKHOLD U, POMMERENINGRÖSER A, JURETSCHKO S, SCHMID M C, KOOPS H P, WAGNER M. Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis: implications for molecular diversity surveys., 2000, 66(12): 5368-5382.
[16] MONTEIRO M, SÉNECA J, MAGALHÃES C. The history of aerobic ammonia oxidizers: from the first discoveries to today., 2014, 52(7): 537-547.
[17] TREUSCH A, LEININGER S, KLETZIN A, SCHUSTER S, KLENK H, SCHLEPER C. Novel genes for nitrite reductase and Amo-related proteins indicate a role of uncultivated mesophilic crenarchaeota in nitrogen cycling., 2005, 7(12): 1985-1995.
[18] BROCHIERARMANET C, BOUSSAU B, GRIBALDO S, FORTERRE P. Mesophilic crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota., 2008, 6(3): 245-252.
[19] PESTER M, SCHLEPER C, WAGNER M. The Thaumarchaeota: an emerging view of their phylogeny and ecophysiology., 2011, 14(3): 300-306.
[20] SINGH B P, HATTON B J, BALWANT S, COWIE A L, KATHURIA A. Influence of biochars on nitrous oxide emission and nitrogen leaching from two contrasting soils., 2010, 39(4): 1224-1235.
[21] STEINER C, TEIXEIRA W G, LEHMANN J, NEHLS T, DE MACÊDO J L V, BLUM W E H, ZECH W. Long term effects of manure, charcoal and mineral fertilization on crop production and fertility on a highly weathered Central Amazonian upland soil., 2007, 291(1/2): 275-290.
[22] RONDON M A, LEHMANN J, RAMÍREZ J, HURTADO M. Biological nitrogen fixation by common beans (L.) increases with bio-char additions., 2007, 43(6): 699-708.
[23] BALL P N, MACKENZIE M D, DELUCA T H, HOLBEN W E. Wildfire and charcoal enhance nitrification and ammonium-oxidizing bacterial abundance in dry montane forest soils., 2010, 39(4): 1243-1253.
[24] SPOKAS K A, REICOSKY D C. Impacts of sixteen different biochars on soil greenhouse gas production., 2009, 3(1): 179-193.
[25] ABUJABHAH I S, DOYLE R B, BOUND S A, BOWMAN J P. Assessment of bacterial community composition, methanotrophic and nitrogen-cycling bacteria in three soils with different biochar application rates., 2018, 18(1): 148-158.
[26] LIU S N, MENG J, JIANG L L, YANG X, LAN Y, CHENG X Y, CHEN W F. Rice husk biochar impacts soil phosphorous availability, phosphatase activities and bacterial community characteristics in three different soil types., 2017, 116: 12-22.
[27] MAGOČ T, SALZBERG S L. FLASH: fast length adjustment of short reads to improve genome assemblies., 2011, 27(22): 2957-2963.
[28] EDGAR R C. UPARSE: highly accurate OTU sequences from microbial amplicon reads., 2013, 10(10): 996.
[29] WANG Q, GARRITY G M, TIEDJ J M, COLE J R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy., 2007, 73(16): 5261-5267.
[30] CHAO A, BUNGE J. Estimating the number of species in a stochastic abundance model., 2002, 58(3): 531-539.
[31] LANGILLE M G I, ZANEVELD J, CAPORASO J G, McDonald D, Knights D, Reyes J A, Clemente J C, Burkepile D E, Thurber R L V, Knight R, Beiko R G, Huttenhower C. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences., 2013, 31(9): 814.
[32] 郭赟. 长期施肥对酸性及中性水稻土壤中氨氧化微生物的影响[D]. 南京: 南京师范大学, 2013.
GUO Y. Effects of long-term fertilization on ammonia-oxidizing microorganisms in acidic and neutral rice soils[D]. Nanjing: Nanjing Normal University, 2013. (in Chinese)
[33] PROSSER J I. Autotrophic nitrification in bacteria., 1989, 30(1): 125-181.
[34] LEININGER S, URICH T, SCHLOTER M, SCHWARK L, QI J, NICOL G W, PROSSER J I, SCHUSTER S C, SCHLEPER C. Archaea predominate among ammonia-oxidizing prokaryotes in soils., 2006, 442: 806-809.
[35] NICOLE G W, LEININGER S, SCHLEPER C, PROSSER J I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria.2010, 10(11): 2966-2978.
[36] NEJC S, GUBRYRANGIN C, ŠPELA H, GRAEME W N, INES M, JAMES I P. Thaumarchaeal ammonia oxidation in an acidic forest peat soil is not influenced by ammonium amendment., 2010, 76(22): 7626.
[37] LIAO X, CHEN C, ZHANG J, DAI Y, ZHANG X, XIE S. Operational performance, biomass and microbial community structure: impacts of backwashing on drinking water biofilter.l, 2015, 22(1): 546.
[38] HE L L, BI Y C, ZHAO J, PITTELKOW C M, ZHAO X, WANG S Q, XING G X. Population and community structure shifts of ammonia oxidizers after four-year successive biochar application to agricultural acidic and alkaline soils., 2018, 619: 1105-1115.
[39] 李双双, 陈晨, 段鹏鹏, 许欣, 熊正琴. 生物质炭对酸性菜地土壤NO排放及相关功能基因丰度的影响. 植物营养与肥料学报, 2018, 24(2): 414-423.
LI S S, CHEN C, DUAN P P, XU X, XIONG Z Q. Effects of biochar on NO emission and related functional gene abundance in acidic vegetable soils., 2018, 24(2): 414-423. (in Chinese)
[40] 刘远, 朱继荣, 吴雨晨, 束良佐. 施用生物质炭对采煤塌陷区土壤氨氧化微生物丰度和群落结构的影响.应用生态学报, 2017, 28(10): 3417-3423.
LIU Y, ZHU J R, WU Y C, SHU L Z. Effects of application of biomass carbon on ammonia microbial abundance and community structure in coal mining subsidence area.2017, 28(10): 3417-3423. (in Chinese)
[41] BI Q F, CHEN Q H, YANG X R, LI H, ZHENG B X, ZHOU W W, LIU X X, DAI P B, LI K J, LIN X Y. Effects of combined application of nitrogen fertilizer and biochar on the nitrification and ammonia oxidizers in an intensive vegetable soil., 2017, 7: 108.
[42] WU H P, ZENG G M, LIANG J, CHEN J, XU J J, DAI J, LI X D, CHEN M, XU P, ZHOU Y Y, Li F, HU L, WAN J. Responses of bacterial community and functional marker genes of nitrogen cycling to biochar, compost and combined amendments in soil.2016, 100: 8583-8591.
[43] JIN H. Thesis: Characterization of microbial life colonizing biochar and biochar-amended soils[D]. Ithaca: Cornell University, 2010.
[44] KHODADAD C L M, ZIMMERMAN A R, GREEN S J, UTHANDI S, FOSTER J S. Taxa-specific changes in soil microbial community composition induced by pyrogenic carbon amendments.2011, 43(2): 385-392.
[45] ALLISON S D, MARTINY J B H. Resistance, resilience, and redundancy in microbial communities., 2008, 105(32): 11512-11519.
[46] BÖRJESSON G, MENICHETTI L, KIRCHMANN H, KATTERER T. Soil microbial community structure affected by 53 years of nitrogen fertilisation and different organic amendments., 2012, 48(3): 245-257.
[47] HALLIN S, JONES C M, SCHLOTER M, PHILIPPOT L. Relationship between N-cycling communities and ecosystem functioning in a 50-year-old fertilization experiment., 2009, 3(5): 597.
[48] HARTMANN M, FLIESSBACH A, OBERHOLZER H R, WIDMER F. Ranking the magnitude of crop and farming system effects on soil microbial biomass and genetic structure of bacterial communities., 2006, 57(3): 378-388.
[49] ZHONG W H, GU T, WEI W, ZHANG B, LIN X, HUANG Q, SHEN W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity., 2010, 326(1/2): 511-522.
[50] GEISSELER D, LAZICKI P A, SCOW K M. Mineral nitrogen input decreases microbial biomass in soils under grasslands but not annual crops.2016, 106: 1-10.
[51] OGILVIE L A, HIRSCH P R, JOHNSTON A W B. Bacterial diversity of the Broadbalk ‘Classical’ winter wheat experiment in relation to long-term fertilizer inputs., 2008, 56(3): 525-537.
Short-term Effect of Biochar Amendments on Total Bacteria and Ammonia Oxidizers Communities in Different Type Soils
ZHANG MengYang1, XIA Hao1, LÜ Bo1, CONG Ming1, SONG WenQun2, JIANG CunCang1
(1Microelement Research Center, College of Resources and Environment, Huazhong Agricultural University, Wuhan 430070;2Xinfeng’s Agricultural Technology Promotion Station, Ganzhou 341600, Jiangxi)
【Objective】Ammonia oxidation is the first step in nitrification and the rate-limiting step in nitrification. It is a key link in the global nitrogen cycle. The purpose of this experiment was to study the effects of peanut shell biochar application on bacteria and ammonia oxidation in different type soils in China, so as to provide a theoretical basis for the promotion and use of biochar.【Method】Yellow-brown soil, fluvo-aquic soil and black soil were utilized as the tested soil. Through short-term culture experiments, 16SrRNA sequencing was used to study the effects of biochar on ammonia-oxidizing microorganisms, bacterial community structure and related enzyme gene expression in different type soils. Four treatments for each soil included CK (no fertilizer and biochar), F (single fertilization), C (single 2% peanut shell biochar), and FC (application of fertilizer + 2% peanut shell biochar).【Result】Acid soil pH increased significantly by 0.5-1.0 units after application of biochar (C, FC), but alkaline soil pH decreased significantly by 0.5-0.6 units; the microbial abundance and diversity of yellow-brown soil increased significantly caused by single application of biochar (C). The fluvo-aquic only significantly increased the microbial diversity index of the soil when it was applied to biochar alone (C). Biochar and chemical fertilizers did not significantly change soil abundance and diversity in black soil; the abundance of ammonia-oxidizing bacteria in three soils was higher than that of ammonia-oxidizing archaea. The measured OTU abundance of ammonia-oxidizing bacteria was about 8.1 times that of ammonia-oxidizing archaea. Biochar and chemical fertilizers did not significantly alter the OTU abundance in the thaumarchaeota, but had a significant effect on the OTU abundance in the beta and gamma proteobacteria. The ammonia-oxidizing bacteria of the three soils were mainly β-proteobacteria, accounting for about 60%. In addition, the application of biochar (C, FC) significantly changed the microbial community structure of yellow-brown soil on PC1 (40.4%), and significantly changed the microbial community structure of fluvo-aquic soil on both PC1 (42.3%) and PC2 (21.3%). After application of biochar (C, FC), the expression of ammonia synthesis related enzyme gene in fluvo-aquic soil decreased significantly by 14.7%-39.9% in a short period of time, and the ammoxidation archaea abundance decreased by 70.5% and 48.7% under C treatment and FC treatment, respectively.【Conclusion】After application of biochar, the microbial community structure of yellow-brown soil and fluvo-aquic soil was significantly changed, and the ammoxidation of fluvo-aquic soil was obviously inhibited in a short period of time.
biochar; chemical fertilizer; ammonia oxidation; microbial community structure; yellow-brown soil; fluvo-aquic soil; black soil
10.3864/j.issn.0578-1752.2019.07.013
2018-09-10;
2018-10-29
国家重点研发计划(2017YFD0200800)
张梦阳,Tel:027-87287141;E-mail:zhangmengyang@webmail.hzau.edu.cn。通信作者姜存仓,Tel:027-87287141;E-mail:jcc2000@mail. hzau.edu.cn
(责任编辑 李云霞)
