不同生境间龟鳖类体型的差异
2019-04-11肖繁荣汪继超史海涛
肖繁荣, 汪继超, 史海涛
(热带岛屿生态学教育部重点实验室,海南师范大学生命科学学院,海口571158)
体型(即个体大小)是动物重要的形态特征,它对动物的生境利用、能量需求和生活史有着重要的影响(Cohenetal.,1993;Principe,2008)。动物体型与生境的关系在生态学和进化生物学领域备受关注(Linzmeier & Ribeiro-Costa,2011)。在诸多脊椎动物类群中,体型与家域正相关(Tershy,1992;Pyron,1999;Perry & Garland,2002)。鳄类中的大体型物种通常栖息在缺少水草的开阔生境,而小体型物种生活在热带森林的小溪流中(Farlow & Pianka,2002);占据开阔地面生境的蜥蜴物种体型较大,而占据岩石生境的物种体型较小,树丛物种的体型则介于两者之间(Collaretal.,2011)。
龟鳖类占据海洋、岛屿以及大陆的陆地和淡水等多样化生境,且体型变异大,从背甲长为10 cm的斑点陆龟Homopussignata到背甲长达200 cm的棱皮龟Dermochelyscoriacea(Ernst & Lovich,2009)。这种体型差异主要受捕食压力(Arnold,1979)、种间竞争压力(Vermeij,1994)、迁徙选择(Luschietal.,1998)、食物资源(Headetal.,2009)、生境的物理条件(Bondi & Marks,2013)和气候条件(龟鳖类贝格曼规律;Angielczyketal.,2015)等生态因素的影响。因此,龟鳖类体型与生境存在一定的相关性。
Jaffe等(2011)研究龟鳖类体型与生境之间的关系发现,从海洋到岛屿性陆地,再到大陆性陆地和淡水生境,龟鳖类体型有逐渐变小的趋势。然而,大陆的陆地生态系统和淡水生态系统比单一的海洋或岛屿生境更复杂,如陆地生态系统包含高原、平原和荒漠等生境,淡水生态系统包含不同面积和水流速度的江河、湖泊、池塘以及山间溪流等生境。由于Jaffe等(2011)的研究主要是阐述岛屿性陆龟巨型化的原因,未对大陆陆地生态系统和淡水生态系统进行详细划分,因此尚不清楚这2种生态系统的不同生境中龟鳖类体型是否存在差异。该研究涉及226种,仅占龟鳖目Tesudines总物种数(335种;van Dijketal.,2014)的67.5%,没有涵盖所有龟鳖类物种。本文通过收集331种龟鳖类的形态数据和生境信息,将生境分为海洋、淡水、岛屿性陆地和大陆性陆地4种类型,并将淡水生境分为5种亚类型,大陆性陆地生境分为3种亚类型,从而比较不同生境类型或亚类型之间龟鳖类体型的差异,以揭示龟鳖类体型与生境之间的关系,为龟鳖类的保护提供科学依据。
1 材料与方法
1.1 数据收集
本文使用龟鳖类形态指标中最常用、最稳定和最容易获得的指标——背甲长表示龟鳖类体型。参考《世界龟鳖志》(Ernst & Barbour,1989;Boninetal.,2006)和《中国动物志 爬行纲》(张孟闻等,1998)等资料,获取每一个物种的最大背甲长。由于许多龟鳖类的体型存在两性异形现象(Berry & Shine,1980;Stephens & Wiens,2009),根据Jaffe等(2011)和Angielczyk等(2015)的方法,本文收集文献中该物种最大的背甲长作为其体型指标。共收集了331种龟鳖类的体型数据,包括了龟鳖目98.8%的物种。
1.2 生境分类
龟鳖类研究中常见的生境分类方法主要有5种:1)陆生和水生生境2种类型(Claudeetal.,2003,2004);2)陆生、水生和半水生生境3种类型(Bensonetal.,2011);3)陆生、水生、底栖和海洋生境4种类型(Munteanu,2014);4)海洋、岛屿、淡水和大陆性陆生生境4种类型(Jaffeetal.,2011;Angielczyketal.,2015);5)所有水域、大流水水域、小静水水域、完全陆生、陆生时常下水、陆生偶尔下水6种生境类型(Joyce & Gauthier,2004)。本研究结合上述分类方法,根据龟鳖类生境特征对其生境进行分类。参照Jaffe等(2011)和Angielczyk等(2015)的方法将龟鳖类生境分为淡水、海洋、岛屿性陆地和大陆性陆地4种类型。在Joyce和Gauthier(2004)的基础上,根据水域面积和水流速度将淡水生境分为大流水、大静水、小流水、小静水和所有水域5种亚类型。根据地形地势和干旱程度将大陆性陆地划分为平地、高地和荒漠3种亚类型。根据每一个物种的生境偏好,将其划分到上述4种类型或8种亚类型中(表1)。
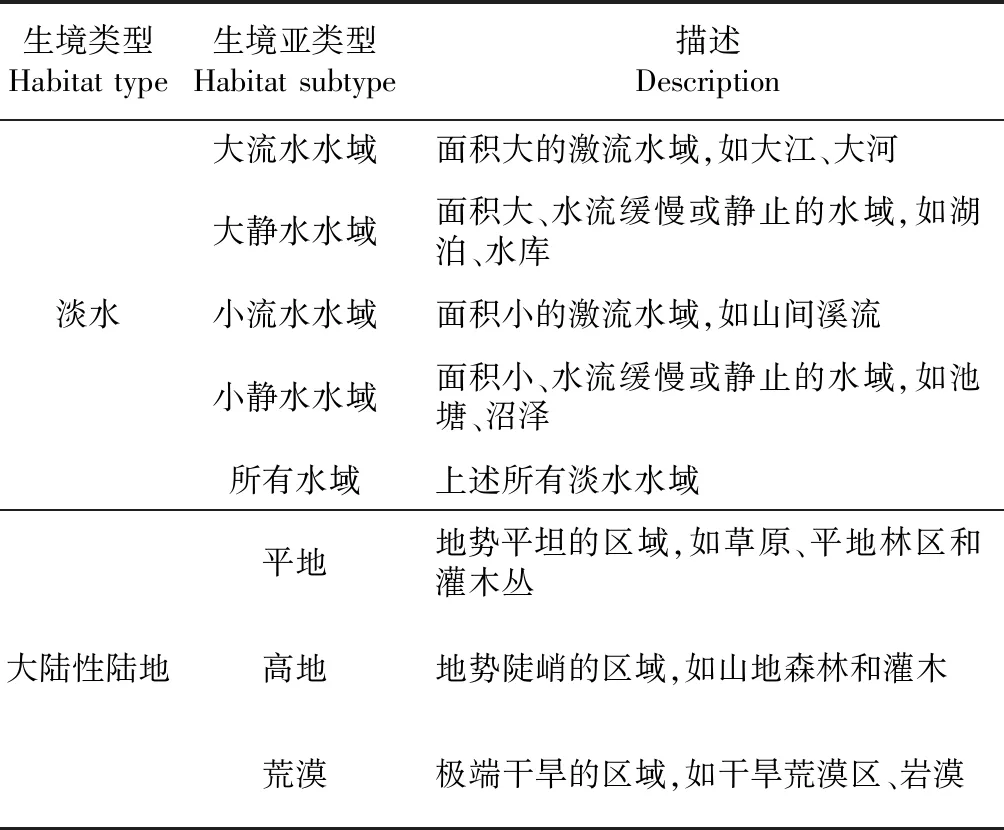
表1 淡水和大陆性陆地的生境分类Table 1 Habitat categories of freshwater and mainland
1.3 数据分析
数据的统计分析和作图分别使用SPSS 19.0和SigmaPlot 12.5完成。统计分析前对数据作对数转换,转换后的数据用于后续的统计检验。采用Kolmogorov-Smirnov检验分析所有龟鳖类以及淡水龟鳖和大陆性陆龟的体型是否符合正态分布,并作频数分布直方图。为消除系统发生对生境的影响,使用以生境类型或亚类型为固定变量,科、属(龟鳖类共14科94属;van Dijketal.,2014)为随机变量的广义线性混合模型(generalized linear mixed model,GLMM)分析龟鳖类体型在不同生境中是否存在差异,并用Bonferroni法进行多重比较。原始数据以平均值±标准差(Mean±SD)表示,P<0.05即认为差异有统计学意义。
2 结果
所有龟鳖类体型的Kolmogorov-Smirnov检验和直方图表明其分布不符合正态分布(n=331,Z=1.953,P=0.001;图1),其中,海龟(n=7,Z=0.693,P=0.723)和岛屿性陆龟(n=21,Z=0.740,P=0.645)的体型符合正态分布。GLMM分析结果显示,消除系统发生的影响后,不同生境类型之间的龟鳖类体型间的差异有统计学意义(F=21.675,P<0.000 1;图2),其中,海龟体型最大(134.86 cm±70.14 cm),显著大于岛屿性陆龟(80.05 cm±37.11 cm;t=4.210,P<0.000 1),岛屿性陆龟体型显著大于淡水龟鳖(32.60 cm±20.57 cm;t=3.633,P=0.001)和大陆性陆龟(27.08 cm±15.77 cm;t=5.574,P<0.000 1),但淡水龟鳖和大陆性陆龟的差异无统计学意义(t=1.743,P=0.082)。
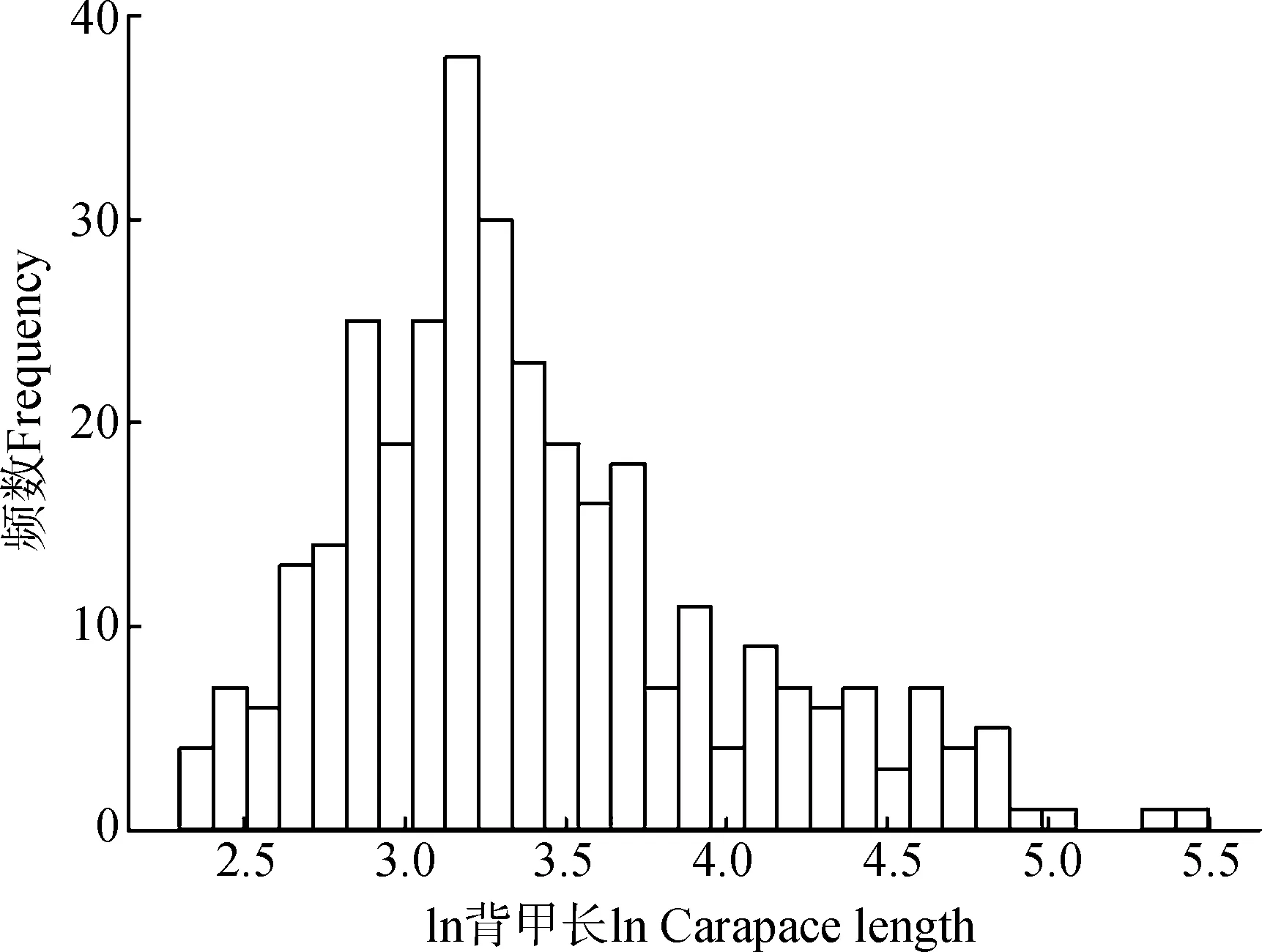
图1 龟鳖类体型分布Fig. 1 Distribution of body size of turtles

图2 不同生境类型中龟鳖类体型的差异Fig. 2 Differences of body size among habitat types for turtles
淡水龟鳖的体型符合正态分布(n=247,Z=1.342,P=0.055;图3)。GLMM结果显示,淡水龟鳖体型在5种生境亚类型之间的差异有统计学意义(F=12.133,P<0.000 1;图4),其中,大静水水域的体型(50.30 cm±27.03 cm)显著大于小静水(23.78 cm±7.47 cm;t=6.101,P<0.000 1)和小流水水域(23.79 cm±10.57 cm;t=5.776,P<0.000 1),大流水水域的体型(39.86 cm±20.34 cm)显著大于小静水水域(t=2.943,P=0.025)和小流水水域(t=3.541,P=0.004),而大静水与大流水水域(t=2.001,P=0.233)、小静水与小流水水域(t=1.172,P=0.485)之间的差异无统计学意义(图4)。这表明水域面积大的体型大,面积小的体型小,而静水、流水对体型无影响。广布所有淡水水域的龟鳖体型(41.50 cm±32.65 cm)比大静水(t=-1.837,P=0.269)和大流水水域(t=-0.486,P=0.627)的小,而比小静水(t=1.631,P=0.312)和小流水水域(t=2.288,P=0.138)的大,趋于中间类型,但差异无统计学意义(图4)。
大陆性陆龟的体型符合正态分布(n=56,Z=0.700,P=0.712)。大陆性陆龟的体型从高地(24.86 cm±10.09 cm)到平地(26.15 cm±20.12 cm)再到荒漠(39.11 cm±19.43 cm)生境有逐渐变大的趋势,但差异无统计学意义(F=0.238,P=0.789)。
3 讨论
本研究表明,海龟体型最大,岛屿性陆龟次之,淡水龟和大陆性陆龟体型最小。这与Jaffe等(2011)的研究结论一致。海龟比其他龟鳖类体型大,主要有4个原因:1)食物链随着海洋初级生产者的数量和营养质量增加而变长,从而导致高营养级的动物进化出较大的体型(Bambach,1993;Heimetal.,2015)。2)海龟较大的体型有利于降低被大型捕食者捕食的风险。晚中生代,海洋中有大量大型沧龙类和上龙类捕食者,在这种捕食压力下,进化出了最大体型的海龟物种(Bernardetal.,2010;Motani,2010)。到新生代,较小的鲨鱼和齿鲸类成为海龟的主要捕食者,海龟体型有所减小(Lutz & Musick,1997)。然而,由于现存海洋捕食者均比其陆地的同类物种大,现存海龟依然维持较大的体型(Jaffeetal.,2011)。3)较大的体型有利于海龟季节性的长距离迁徙,如绿海龟Cheloniamydas能从大西洋的阿森松岛迁徙2 300 km到达巴西海岸(Luschietal.,1998)。4)海龟分布区域广,从赤道到南部或北部均有分布,不同区域之间的海水温差较大,较大的体型能降低表面积与体积比以减少热量散失,从而使海龟适应低温区域的生境(Mrosovsky,1980)。

图3 淡水龟鳖类体型分布Fig. 3 Distribution of body size of freshwater turtles
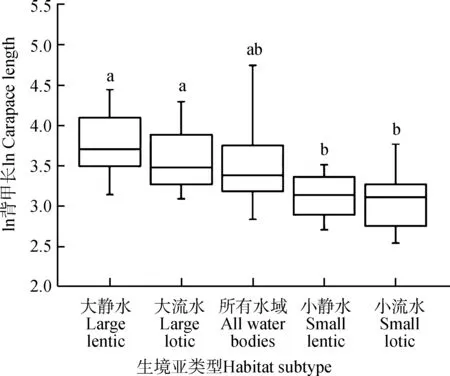
图4 不同生境亚类型中淡水龟鳖类体型的差异Fig. 4 Differences of body size among habitat subtypes for freshwater turtles
岛屿性陆龟的体型显著大于大陆性陆龟,这与Lomolino(2005)和Jaffe等(2011)研究一致。岛屿效应理论认为,受岛屿捕食压力、种间竞争压力、食物资源和迁移选择等因素的影响,岛屿动物常向巨型化或侏儒化方向进化(van Valen,1973;Lomolino,1985)。岛屿效应已在两栖爬行类、鸟类和哺乳类上得到验证(Lomolino,2005;Lietal.,2011)。岛屿上天敌数量少,种内竞争是其主要的选择压力,大体型在种内竞争占优势,从而导致岛屿物种朝巨型化方向进化(Palkovacs,2003)。岛屿性陆龟巨型化还是一种加强扩散能力的预适应,有利于祖先种群在岛屿上的早期定殖,从而促进巨型化特征稳定地遗传到后代个体中(Pritchard,1996)。同时,缺乏食草动物的岛屿环境也是导致岛屿性陆龟巨型化的重要因素,因为岛屿性陆龟为草食性动物,缺乏食性相同的竞争者(Arnold,1979)。此外,巨型化岛屿性陆龟的禁食能力较强,干旱时期有利于它们在取食场和水源地之间进行长距离的迁移(Arnold,1979;Lomolino,2005)。
大陆性陆龟与淡水龟鳖的体型之间的差异无统计学意义,这可能是水陆环境的差异对体型影响较小,而对身体形状(背甲高与背甲宽的比值)影响较大的缘故,研究表明陆生龟类的背甲高拱,而水生的背甲比较扁平,呈流线型(Romer,1967;Bonnetetal.,2010;Bensonetal.,2011)。淡水龟鳖和大陆性陆龟占据从干旱荒漠到温带森林的多种水域和陆地生境,这些复杂生境中多种生态因子的综合作用也可能导致它们体型相似(Jaffeetal.,2011)。但大陆性陆龟或淡水龟鳖的多样化生境与较宽的体型之间可能存在一定关联。本研究发现大陆性陆龟的体型从高地到平地再到荒漠生境有逐渐变大的趋势。合理的解释是生活在地势陡峭岩石生境(高地)的物种需要攀爬,较小的体型有利于其灵活运动,也有利于其寻找诸如石缝这样狭小的隐蔽场所,如平顶闭壳龟Cuoramouhotii和扁平陆龟Malacochersustornieri,而在开阔平地的陆龟则无此需求,因此体型较大(Ireland & Gans,1972;Collaretal.,2011;Xiaoetal.,2017)。荒漠生境中的陆龟体型比其他2种生境的大,这可能与岛屿性陆龟一样,较大的体型有利于它们在干旱的环境中长距离迁移寻找食物和水源(Arnold,1979;Lomolino,2005)。但这种逐渐变大的趋势并不显著,可能是由于不同生境中的一些陆龟具有相同的生活习性,从而具有相似体型。如栖息于海岸线附近(平地)的纳米比亚珍龟Homopussolus和山区的扁平陆龟为适应穴居生活,体型均较小(Malonza,2003;Boninetal.,2006)。
进一步比较淡水龟鳖在5种不同生境亚类型中的体型,发现淡水龟鳖体型与水域面积相关,水域面积大的体型大,面积小的体型小,而体型与水域是静水或流水无关。面积大的水域能承载大体型的龟鳖,并能为其提供丰富的食物资源。如黄斑巨鳖Rafetusswinhoei和鼋Pelochelyscantorii栖息在面积较大的湖泊和江河中(张孟闻等,1998),其体型显著比其他淡水龟鳖大。但分布在山间溪流、池塘等小水域的龟鳖,由于受水流量和水中食物资源的限制,生长缓慢,体型偏小。Bondi和Marks(2013)研究石斑水龟Actinemysmarmorata不同种群的体型也发现相似的规律,栖息在水流量和面积较大水域的个体比在水流量和面积较小水域的个体大。此外,与海洋环境类似,淡水脊椎动物与捕食者的体型是协同进化的关系,为防止被捕食,淡水脊椎动物进化出较大的体型(Perssonetal.,1996;Gaetaetal.,2018)。因此,淡水龟鳖类在面积大的水域体型较大可能与该水域现存或历史上存在大型捕食者有关。相对而言,小水域的淡水龟鳖类个体小,一方面由于其捕食者体型小,另一方面则是因为小个体的淡水龟鳖类灵活度高,掉头转身的速度快,从而能有效地逃避捕食者(Fish & Nicastro,2003;Riveraetal.,2006)。已有研究表明,流水水域的优雅伪龟Pseudemysconcinna种群比静水水域的种群更呈流线型(Rivera,2008)。因此,静水或流水可能只作用在淡水龟鳖的身体形状上,而对其体型无影响,从而导致同一大小水域中流水与静水生境的龟鳖体型无显著差异。广布所有水域的龟鳖体型趋于中间型,这是为兼顾适应不同特征的淡水生境,对这些生境的利用存在权衡的结果。
本研究首次发现淡水龟鳖类的体型与水域面积有关,水域面积越大,其体型越大。这对珍稀大型淡水龟鳖类的保护具有重要的实践指导意义。如,黄斑巨鳖历史上曾经分布于我国黄河、长江、太湖和钱塘江等流域,但人类活动导致动物的栖息地丧失和破碎化,使得这些大型江河或湖泊被隔离,水域面积缩小,而该物种又不能在短时间内进化出较小体型以适应破碎的小水域,加之其他因素导致这些流域的野生个体灭绝。现存于我国境内红河流域的黄斑巨鳖,近年来由于干流梯级水电开发密度急剧下降,野生种群濒临灭绝(王剑,史海涛,2011)。本研究表明龟鳖类根据其体型适应特定的生境,大体型的淡水龟鳖占据大的水域,当其所依赖的大型水域遭到破坏,其生存将会受到影响,这是黄斑巨鳖等大型淡水龟鳖类濒临灭绝的重要原因之一。因此,要保护这些大型珍稀龟鳖类就要保护其所依赖的淡水生态系统的完整性。
致谢:墨西哥国立自治大学(Universidad Nacional Autónoma de México)的Taggert Butterfield博士研究生对英文摘要的修改,谨致诚挚谢意。
