雄性峨眉髭蟾“胡子”的组织结构特征
2019-04-11张武元陆宇燕史静耸朱磊李丕鹏
张武元, 陆宇燕, 史静耸, 朱磊, 李丕鹏*
(1.沈阳师范大学两栖爬行动物研究所,沈阳110034; 2. 沈阳师范大学辽宁省生物进化与多样性重点实验室,沈阳110034;3.沈阳师范大学生命科学学院,沈阳110034; 4. 中国科学院古脊椎动物与古人类研究所,北京100044;5.中国科学院脊椎动物演化与人类起源重点实验室,北京100044; 6. 成都观鸟会,成都610041)
峨眉髭蟾Leptobrachiumboringii(Liu, 1945)隶属角蟾科Megophryidae拟髭蟾属Leptobrachium(Zhengetal.,2008),俗称“胡子蛙”或“角怪”,中国特有种。由于栖息环境质量下降,峨眉髭蟾的种群数量已经非常稀少,世界自然保护联盟(IUCN)红色名录将其列为濒危(EN)物种(Fei & Wu,2004;Fei & Ye,2016)。雄性成体峨眉髭蟾在每年2—4月的繁殖季节出现明显的性二态特征:上颌有10~16枚平均长度为3~5 mm的锥形黑色角质刺,前臂变粗壮,背部及大腿等部位的皮肤变厚、松弛(Zhengetal.,2010;樊均德等,2011)。研究认为,雄性成体峨眉髭蟾上颌角质刺是争夺领地的格斗武器(Hudsonetal.,2010;Hudson & Fu,2013),目前未见角质刺组织学结构的报道。本研究首次对峨眉髭蟾角质刺进行了组织学观察,从而弥补了该物种独特性二态的生物学资料,旨在丰富我国特有无尾两栖类的组织学资料,为进一步对两栖动物进化、行为等方面关系的研究积累科学依据。
1 材料和方法
2016年2月采自四川省峨眉山的3只雄性成体峨眉髭蟾(表1),双毁髓法处死后,10%中性福尔马林溶液固定,每只随机选取2枚大小相似的角质刺,并将其底部相连接的上颌皮肤与上颌骨一同剪下,剪取的样品约0.5 cm×0.5 cm,分别用于常规组织学观察和表面结构的亚显微结构观察。常规石蜡切片之前,用一定浓度的HNO3溶液进行脱钙处理。切片厚度为7 μm,采用苏木精-伊红(HE)染色。利用Olympus BX51-DP71显微镜观察和拍照,用Image-Pro Express 6.0测量单侧角质刺的厚度及表皮嵴长度等,采用SPSS 20处理数据。利用台式电镜HITACHI TM3030观察和拍照。
2 结果
2.1 角质刺结构
光镜下,峨眉髭蟾的角质刺整体呈倒“V”形(图版Ⅰ:1),与皮肤相连接的角质刺单侧厚度为145 μm±10 μm,并可见明暗相间的黑色斑状结构(图版Ⅰ:2)。在高倍物镜下可见角质刺侧壁由排列紧密的角质化上皮细胞构成(图版Ⅰ:3),这些上皮细胞由于胞质着色深浅的差异形成了低倍镜下明暗相间的条带结构。整体上,着色浅部分的上皮细胞胞核淡染,呈瘦弱的梭形;胞质有一定的膨胀现象,着色极浅;细胞间质明显,呈强噬伊红。角质刺深染部分,胞核依稀可见,呈椭圆形;胞质呈深棕色;界限基本清晰,细胞排列紧密(图版Ⅰ:4)。在角质刺内侧可清晰分辨皮肤的表皮层和真皮层,但在表皮和与角质刺内侧的接触面仅可见不同程度角质化的细胞,未见其他结构填充(图版Ⅰ:5)。在角质刺基部可见明显的皮肤褶翻起将其包裹在内(图版Ⅰ:6),并可见皮肤褶向上延伸形成的由单层立方上皮细胞构成的外膜。但由于角质刺较为坚硬,这2种结构在制片过程中大多被破坏而丧失(图版Ⅰ:7)。
扫描电镜下,角质刺顶面观(图版Ⅱ:8)为锥形的“小山丘”状,在其顶部尚可分辨出表皮细胞的轮廓,在其侧面可见呈覆瓦状排列的角质化细胞出现层层堆积的特征(图版Ⅱ:9)。在角质刺和上颌皮肤交界处可见两者结构是一致的,均由多边形的角质化表皮细胞构成,仅在角质刺处出现隆起(图版Ⅱ:10)。构成角质刺的表皮细胞的侧缘具有数个指状突起,与相邻细胞之间形成良好的镶嵌连接。而在表皮细胞的上、下表面上可见大量的突起或凹陷,呈现出凹凸不平的花纹结构,即上、下层表皮细胞之间具有大量的镶嵌连接(图版Ⅱ:11)。

图版Ⅰ 雄性峨眉髭蟾角质刺的显微结构Plate Ⅰ Microstructure of the keratinized nuptial spines of male Leptobrachium boringii

图版Ⅱ 峨眉髭蟾角质刺的扫描电镜图Plate Ⅱ Scanning electron microscopy of the keratinized nuptial spines of Leptobrachium boringii
2.2 角质刺处及其周边的皮肤结构
在倒“V”形角质刺中央的凹陷处,上颌表皮隆起,与角质刺紧密连接,形成了角质刺的“基座”(图版Ⅰ:1、7)。此处仍可区分表皮和真皮,表皮层由内到外可以区分出生发层、棘细胞层、颗粒层和角质层。生发层位于表皮最内层,与真皮层连接,为1层矮柱状细胞,排列紧密,胞质弱酸性;胞核较大、椭圆形,多位于近细胞基底部,深染。棘细胞层细胞多为椭圆形、梭形或不规则形,胞质着色略深,胞核趋向圆形,着色略变浅。颗粒层细胞多为梭形,胞质嗜酸性逐渐增强;细胞核着色不一,逐渐变为梭形,横位(图版Ⅰ:5,图版Ⅲ:12)。最外层为角质层(图版Ⅲ:13),细胞层数1~3,胞质为强嗜酸性,被染成深红色,而胞核着色不明显;角质层下的表皮层由5~11层细胞组成;而在拱起的顶部表皮层多达10~13层细胞(图版Ⅲ:14)。角质刺侧壁表皮层常有1~2个伸入到真皮层深处的表皮嵴(图版Ⅲ:15),其为表皮基底层细胞向真皮层的延伸。角质刺内的真皮层没有典型的疏松层和致密层,纤维成分多呈多向性分布,其间夹杂着疏松结缔组织(图版Ⅲ:16、17),可见毛细血管和色素细胞(图版Ⅲ:18),但未见皮肤腺。在角质刺基部与上颌皮肤连接处以及其周边皮肤结构可见真皮层结构的渐变过程,尤其是周边皮肤的真皮致密层内纤维呈束状,并呈平行分布,而纤维束进入角质刺基部后呈发散状,失去束状形态结构特点(图版Ⅲ:19)。
峨眉髭蟾上颌角质刺周边的皮肤均可分为典型的表皮层和真皮层,真皮层中疏松层与致密层清楚可辨,并可见大量皮肤腺体、色素细胞(图版Ⅲ:20、21)。
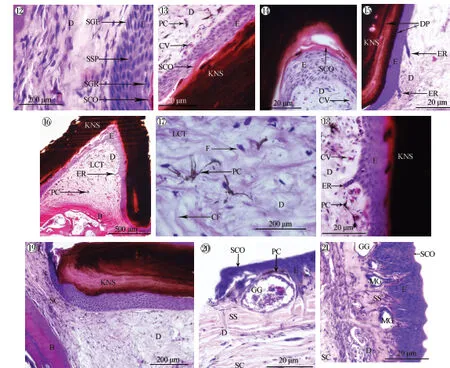
图版Ⅲ 峨眉髭蟾角质刺处及其周边皮肤的显微结构Plate Ⅲ Microstructure of the skin on and around the keratinized nuptial spines of Leptobrachium boringii
3 讨论
3.1 峨眉髭蟾角质刺的结构特征
Liu(1945)依据峨眉髭蟾为模式种建立的髭蟾属Vibrissaphora与拟髭蟾属的最大区别是其雄性上颌长有角质刺。但髭蟾属的有效性存在争议(Dubois,1980;Dubois & Ohler,1998)。形态学研究表明,髭蟾属和拟髭蟾属物种的成体极为相似,但髭蟾属的雄性在繁殖季节上唇缘有黑色锥状角质刺,能显著区别于拟髭蟾属,故应独立为属。因此,角质刺也一直作为鉴定该属物种重要的分类依据之一(刘承钊等,1980;费梁等,1995)。而近年分子系统学研究认为,之前基于形态订立的髭蟾属和拟髭蟾属互相不成单系,建议把髭蟾属并入拟髭蟾属(Rao & Wilkinson,2008;Zhengetal.,2008;Matsuietal.,2010;Chenetal.,2013;Fei & Ye,2016)。
无尾两栖类特有的第二性征是婚瘤,许多种类在繁殖期雄性拇指皮肤局部特化形成突起的婚垫,有些类群也在其他指和(或)胸部形成刺团(Duellman & Trueb,1994;Lunaetal.,2012)。婚垫有3种形态结构类型,其中1种具有婚刺,一般认为构成婚瘤的刺由表皮和真皮组成(Duellman & Trueb,1994;杨纯,2004;Lunaetal.,2018)。但对分布于墨西哥到阿根廷的拉丁美洲雨蛙科Phyllomedusidae(原为雨蛙科Phyllumodesinae亚科)5属26种婚垫的研究表明:婚刺细小,外部形态复杂多样,由表皮层细胞突起构成,且只有最外层表皮细胞角质化,真皮层不参与刺的形成(Julianetal.,2010)。而峨眉髭蟾的角质刺不仅大,由表皮和真皮组成,真皮深入刺的髓部形成锥状突起的髓芯,且表皮的角质化程度高,表皮细胞层数明显多于其周边皮肤的真皮细胞层数,从而形成强硬的大刺。此外,髭蟾角质刺的数量、大小和形态特征具有种间差异,也是物种鉴别特征之一,这点和棘胸蛙Paaspinosa等的婚垫和(或)胸部刺团相似(俞宝根,2010;费梁等,2012)。
牛角是皮肤的同源器官,为圆锥形管状填充结构,主要由角质外壳、内部骨芯和连接两者的软组织层3部分组成(邹猛等,2015;佘平昌,2016)。在白犀牛Ceratotheriumsimum鼻角观察到宽度为5~6 cm周期性深浅不一的条带状结构,而这些条带正好与其年龄基本一致(Pienaaretal.,1991;Rachlow & Berger,1997;Hieronymusetal.,2006)。峨眉髭蟾上颌角质刺无内部骨芯支持,仅有隆起皮肤作为支撑,同时角质刺处的表皮细胞角质化堆积后直接形成了强大的角质刺结构,两者之间未见其他的组织作为间接介质,这与牛角结构区别明显。雄性成体峨眉髭蟾的角质刺在繁殖季节后期逐个脱落,这与白犀牛的鼻角明显不同。而峨眉髭蟾的繁殖季节是冬春接替的2月下旬至3月中旬,气温波动较大,温度变化直接影响了峨眉髭蟾的各项生理机能,包括皮肤的代谢功能和生长速度。而这种变化是否是导致峨眉髭蟾上颌角质刺切面呈现规律的明暗相间结构的成因还有待进一步积累资料证明。
3.2 角质刺结构与功能的适应性
在无尾两栖类中,仅5%物种的雄性之间为竞争领地和配偶存在着直接格斗行为,但一般均以摔跤的形式进行(Shine,1979;Wells,2007)。而性成熟的雄性峨眉髭蟾却演化出了具有明显格斗功能的第二性征——上颌角质刺,且在繁殖季节能助力于领地和配偶的竞争。因此,Hudson等(2010)认为峨眉髭蟾是研究无尾两栖类动物雄性格斗武器出现与第二性征之间演变关系的有效模型物种。
两栖类皮肤比较特殊,不仅要完成动物皮肤的基本功能,还在呼吸、生殖和防御等方面起重要作用(Dockray & Hopkins,1975;Sekietal.,1989,1995;Brodieetal.,1990;Hofer & Mokri,2000;Blaustein & Belden,2003;梁刚,王琼霞,2004;刘满樱等,2007;曹燕等,2011;吴文英等,2011;雷飞宙等,2012;贾丽丽等,2013;高凤娟等,2016)。与其他同类物种皮肤结构一致,峨眉髭蟾上颌部由皮肤演化出来的角质刺,在繁殖季节用于争夺领地和交配权。当1只没有繁殖巢穴的雄性峨眉髭蟾接近1只有巢穴的雄性峨眉髭蟾并试图侵占领地时,便会发生格斗行为。巢穴中的个体会尝试将入侵者推出巢穴入口,而入侵者则会通过扭抱并旋转对方来获得领地。这个过程中,一旦1只雄性能用粗壮的前臂成功抱住对手的腰部,它便会用角质刺反复刺击对方的腹部,战斗时间通常会持续2~5 min,直到有一方成功被驱赶至巢外。格斗行为对双方均有伤害甚至由于格斗太过激烈而折断角质刺(Hudsonetal.,2010)。峨眉髭蟾上颌角质刺用于雄性间的格斗,所以须承受巨大的冲击力和牵拉力。本实验结果显示,峨眉髭蟾上颌角质刺很好地适应了格斗需要。首先,角质刺的核心由突起的皮肤构成,且真皮部分以发散状多向性分布排列的纤维成分分散于疏松结缔组织中,即以软组织很好地缓冲了来自外部的冲击力;角质刺基部由皮肤包裹更加巩固了角质刺与上颌皮肤的连接,角质刺侧壁表皮伸入真皮部分的表皮嵴加强了表皮与真皮连接的牢固性。而在以往研究中未见无尾两栖类表皮嵴的报道(Penrose & Ohara,1973;Young,1977;王大江等,1991;Hanetal.,2010;徐平等,2012;邵华信,吕学洗,2014;王溯,陈晓蓉,2014)。另外,在角质刺基部和周边皮肤的真皮致密层的排列可以将角质刺承受的冲击力分散到周围,减轻角质刺的压力,很好地保护了角质刺的一定存在周期,有利于保持繁殖期间所需要的竞争力。
对黄牛Bostaurus角以及穿山甲Manispentadactyla鳞片拉伸断口的亚显微结构观察发现,其细胞为角质化的鳞片状细胞层层堆积组成。这种层状的堆积结构在抵抗裂纹扩展中发挥着很大的作用,可以使裂纹在扩展过程中转向,大大增加裂纹扩展的路径、能量,从而大幅度提高材料的断裂韧性(马云海等,2008;李炳蔚,2010;唐伟文,2015)。另外,峨眉髭蟾角质刺的每个角质化细胞上都具有凹凸不平的花纹结构,这对于角质刺的增韧也具有重要作用。首先,角质化细胞表面凹凸不平的花纹可以加强微观的摩擦自锁,即界面粘结。又由于这些细胞之间存在的镶嵌连接和桥粒结构,当其承受载荷时,横向和纵向为数众多的角质化细胞之间会相互作用,从而造成很大的抵抗破坏能力,可以抵抗外界来自不同方向的牵拉力,是一种有效的增韧机制。这使角质刺在保持结构完整性的基础上,同时还具有良好的力学结构,这种存在于无尾两栖类的“格斗武器”也为今后仿生学提供了更多的资料。
致谢:感谢沈阳师范大学生命科学学院孙琦等同学在实验过程中的帮助,感谢峨眉山黄湾乡大峨村村民骆玉泉在作者采集标本过程中遇到生命危险时提供的救援帮助。
