IFN-γ基因敲除小鼠的繁育及其对TgCatCHn4弓形虫的感染研究
2019-04-11,,,
, , ,
γ干扰素(Interferon gamma,IFN-γ),又名Ⅱ型干扰素或免疫干扰素,主要由单核巨噬细胞、T细胞和NK细胞产生[1]。IFN-γ的主要生物学作用是促进免疫、抗病毒和抗肿瘤[2]等,同时还具有抗寄生虫作用[3]。弓形虫作为食源性寄生原虫,会导致人和动物的流产,畸胎或死胎,对人和动物的健康造成威胁[4-5]。弓形虫基因型丰富多样,表现出不同的遗传多样性和致病性,我国主要流行Chinese1(ToxoDB#9)型弓形虫,不同分离株毒力差异较大,部分Chinese1型弓形虫虫株感染小鼠后呈慢性经过[6-9]。目前基因敲除小鼠用于感染性疾病及其免疫应答的研究较多,但还未见关于IFN-γ-/-小鼠对我国弓形虫虫株的感染情况研究。为获得弓形虫弱毒株的急性动物模型,从美国Jackson实验室购入IFN-γ-/-小鼠冷冻胚胎,经复苏培养及6个月的饲养和繁育,最终成功获得了大量IFN-γ-/-小鼠,并对比研究了IFN-γ-/-小鼠和BALB/c小鼠对TgCatCHn4(Chinese1)弓形虫虫株的感染率及宿主存活情况。
1 材料与方法
1.1试验材料 TgCatCHn4分离于猫组织[9],由河南农业大学兽医病理实验室保存。
1.2试验动物 IFN-γ-/-小鼠胚胎购于美国Jackson实验室(Stock No:002287),经北京协和医院医学实验动物研究所复苏,获得6只小鼠,其中雄性3只(纯合子1只,杂合子2只),雌性3只(纯合子1只,杂合子2只)。另从郑州大学试验动物中心购入BALB/c小鼠40只。
1.3IFN-γ-/-小鼠的饲养、繁育及鉴定 小鼠置于实验室SPF环境中,按照相应饲养标准进行饲养。饲养过程中,每日添加饮水、补充饲料,适时更换垫料。小鼠繁殖时,IFN-γ+/-小鼠以雌雄按3∶1比例合笼饲养。哺乳期满的仔鼠与母鼠分开,雌雄分笼饲养,并打耳标、记录数量。
剪取仔鼠(3周龄)尾尖组织0.5 cm,按照组织DNA提取试剂盒(天根生化科技有限生物,批号:DP304)说明提取尾尖DNA。IFN-γ扩增引物1:5′-CCT TCT ATC GCC TTC TTG ACG-3′,引物2:5′-AGA AGT AAG TGG AAG GGC CCA GAA G-3′,引物3:5′-AGG GAA ACT GGG AGA GGA GAA ATA T-3′。目的片段大小为500 bp的条带为IFN-γ-/-小鼠,210 bp的条带为IFN-γ+/+小鼠,两条210 bp和500 bp的条带为IFN-γ+/-小鼠。
1.4小鼠感染弓形虫及弓形虫负载量 从Vero细胞培养瓶中收集TgCatCHn4弓形虫速殖子,IFN-γ-/-小鼠(n=5)和BALB/c小鼠(n=24)分别经背部皮下注射1 mL弓形虫(104个/mL)。常规饲养,每日观察临床表现、死亡日期并记录。接种后30 DPI(30 days post inoculation,30 DPI)的小鼠,对肺脏及肠系膜淋巴结进行涂片镜检,观察是否含有弓形虫速殖子。存活大于30 DPI的小鼠面静脉采血,采用改良凝集试验(Modified Agglutination Test,MAT)检测血清中弓形虫抗体,检查其感染情况。存活大于30 DPI并自然死亡的小鼠,取脑组织压片镜检观察包囊并计数[10]。
将含有弓形虫包囊的小鼠脑组织或含有速殖子的小鼠肺脏或淋巴结,接种于生长良好的Vero细胞培养瓶中,每周更换两次细胞培养液,并在倒置显微镜下观察速殖子。
1.5统计分析 采用软件Graph Pad Prism 5.0 software (Graphpad Software Inc., San Diego, CA, USA) 对数据进行统计分析。P<0.05为差异有统计学意义。
2 结 果
2.1小鼠繁殖结果及生长情况 经IFN-γ+/-小鼠雌性和雄性交配的方法繁育,成功获得了子代小鼠。IFN-γ+/-小鼠毛发光亮顺滑,四肢有力,尾长,行动灵活;IFN-γ-/-小鼠身形较为瘦小,行动灵活,善跳跃。平均每只IFN-γ+/-母鼠每胎生产10~12只幼鼠,经过6个月的繁育共生产小鼠799只,存活758只,存活率达94.9%。
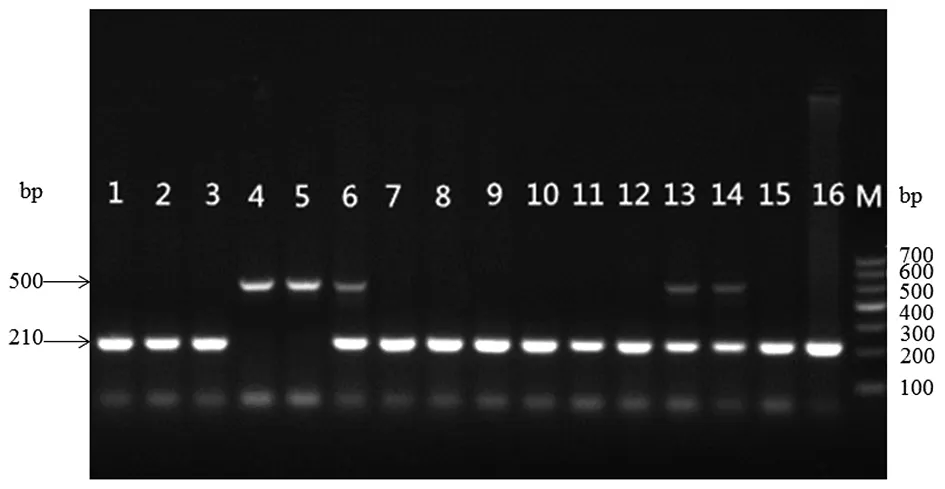
M: DNA Marker 700;1-16:小鼠DNA扩增结果;1-3、7-11、15-16为野生型;4-5为纯合子;6、13-14为杂合子。
2.2子代小鼠IFN-γ基因鉴定结果(图1) 亲代为IFN-γ+/-杂合子小鼠交配,子代小鼠有杂合子、野生型、纯合子3种表现型。使用琼脂糖凝胶电泳进行IFN-γ基因鉴定,引物1和引物3的扩增产物为500 bp大小的片段,引物2和引物3的扩增产物为一条210 bp大小的片段,结果如图1所示。经基因鉴定,获得IFN-γ+/+308只,IFN-γ+/-404只,IFN-γ-/-46只,各种基因所占比例为:IFN-γ+/+占40.6%;杂合子IFN-γ+/-占53.3%;纯合子IFN-γ-/-占6.1%,野生型:杂合子:纯合子的比例约为7∶9∶1。
2.3TgCatCHn4弓形虫对IFN-γ-/-小鼠和BALB/c小鼠的致病性研究(表1) 由表1, IFN-γ-/-小鼠感染104个TgCatCHn4弓形虫速殖子后,存活时间为17.0±1.0 DPI(16DPI~18DPI),急性感染期死亡率为100%,死亡小鼠肺淤血,肺脏涂片可见大量弓形虫速殖子。30 DPI,小鼠血清弓形虫IgG抗体结果显示,BALB/c小鼠的弓形虫感染率为100%,临床表现未见明显异常,急性感染期死亡率为0%,BALB/c小鼠存活时间为304.0±43.7 DPI(51 DPI~726 DPI),自然死亡小鼠只有10%的大脑研磨液中检出弓形虫包囊,包囊数量2.6±1.7个/只小鼠大脑。弓形虫阳性小鼠组织接种Vero细胞后,IFN-γ-/-小鼠肺脏组织和BALB/c小鼠大脑组织内的TgCatCHn4弓形虫在Vero细胞培养后,初次观察到速殖子的时间分别为2 d和19 d。
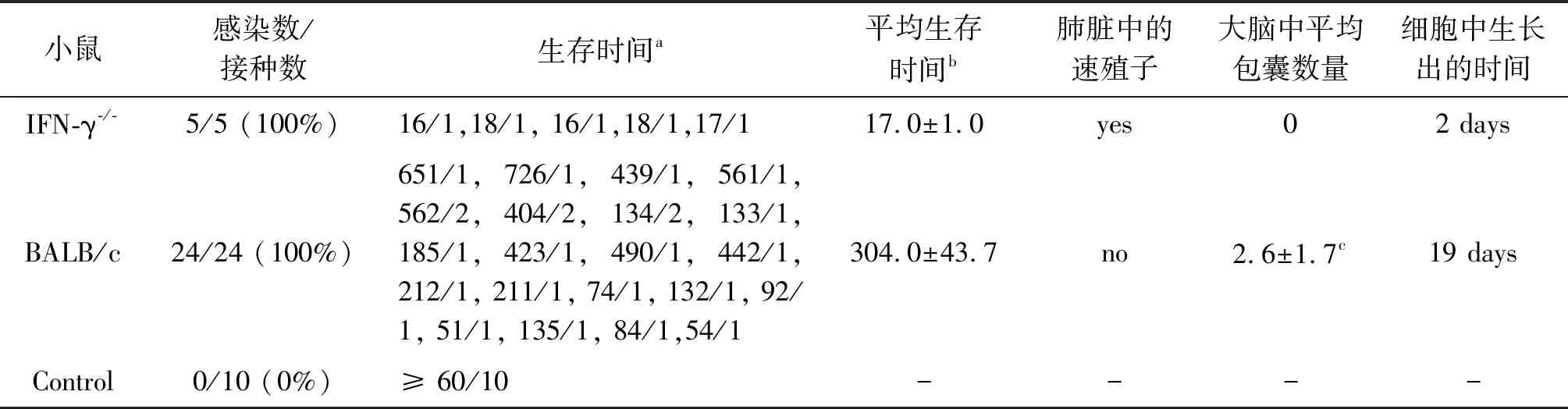
表1 TgCatCHn4株弓形虫对IFN-γ-/-小鼠和BALB/c小鼠的致病性Tab.1 Pathogenicity of TgCatCHn4 Toxoplasma gondii on IFN-γ-/- mice and BALB/c mice
3 讨 论
研究表明,IFN-γ在抗弓形虫感染中起重要作用。体外试验发现,IFN-γ能够抑制弓形虫速殖子繁殖,并可抑制弓形虫包囊活化[11],调控免疫细胞限制弓形虫在大脑的生长[12]。小鼠感染试验也证实,IFN-γ具有抑制弓形虫缓殖子转变为速殖子的作用[13]。
全世界共鉴定了231种弓形虫基因型,1 457株弓形虫虫株(http://ToxoDB.org),我国弓形虫虫株分离较少,目前已从人和动物分离得到约122株弓形虫虫株[9],89%的虫株基因型为ToxoDB#9(Chinese1),Chinese1不同分离虫株间毒力差异较大,部分虫株毒力较弱[6-9],动物模型中常难以收获虫体。可使用猫进行弓形虫卵囊的收获,具有较高的收获率,且耗时短(21 DPI)[9]。因此在弓形虫的致病机制和疫苗研究中,获得弓形虫弱毒株的急性感染模型十分重要。
近年来,有大量试验构建小鼠模型研究弓形虫的致病机理。弓形虫的易感性和致病性与小鼠品系有一定关系,降序排列依次为:IFN-γ基因敲除小鼠、人白细胞抗原(Human Leucocyte Antigen, HLA)A1101转基因小鼠、HLA A0201转基因小鼠、HLA B0702转基因小鼠、Swiss Webster、C57/black、BALB/c[10]。本研究使用IFN-γ-/-小鼠进行弓形虫感染试验研究发现,感染TgCatCHn4弓形虫虫株后,IFN-γ-/-小鼠死亡率100%,均在急性期死亡,肺脏涂片可见大量速殖子阳性,与BALB/c小鼠比较,可在较短时间收获弱毒弓形虫虫体,利于后续研究。本研究结果显示IFN-γ-/-小鼠对弓形虫的易感性强于BALB小鼠,这与报道结果一致。IFN-γ-/-小鼠和BALB/c小鼠分别感染100个Me49株弓形虫卵囊,100% 的IFN-γ-/-小鼠(25/25)呈弓形虫急性感染而死亡,存活期小于14 DPI,但BALB/c小鼠(23/1)的死亡率是4%,大脑平均包囊126个[10]。IFN-γ-/-小鼠和Swiss小鼠感染TgCatCHn1-2,TgCatCZg 1-4后,IFN-γ-/-小鼠均呈弓形虫急性感染,存活期20-27 DPI,而Swiss小鼠未见明显临床症状,存活期超过42DPI[14]。可见,与BALB小鼠和Swiss小鼠比较,IFN-γ-/-小鼠对弓形虫更易感。
对IFN-γ基因敲除小鼠进行繁育,可为饲养繁育该品系小鼠提供技术参考依据。IFN-γ基因敲除小鼠对弓形虫的临床表现,也说明IFN-γ在弓形虫感染过程中具有重要的抗虫免疫作用,为深入研究弓形虫急性感染的免疫机制提供了理想模型。
