防城河口湾潮间带大型底栖动物群落结构研究
2019-04-11赖廷和何斌源黄中坚汤乔覃漉雁朱婷莫珍妮刘俐钟云旭
赖廷和, 何斌源, 黄中坚, 汤乔, 覃漉雁, 朱婷, 莫珍妮,刘俐,钟云旭
防城河口湾潮间带大型底栖动物群落结构研究
赖廷和1, 2, 何斌源1, 2, 黄中坚2, 汤乔3, 覃漉雁2, 朱婷2, 莫珍妮2,刘俐2,钟云旭4
1. 钦州学院, 广西 钦州 535011; 2. 广西壮族自治区海洋研究院, 广西 南宁 530022; 3. 广西北仑河口国家级自然保护区管理处, 广西 防城港 538100; 4. 广西红树林研究中心, 广西 北海 536000
文章根据2016年5月、8月、11月和2017年2月的调查数据, 研究自然和人为因素对广西防城河口湾潮间带大型底栖动物群落结构的影响。结果表明: 4个季度共采集到潮间带大型底栖动物10门252种, 以软体动物门、节肢动物门和环节动物门为主。调查区域的优势种包括珠带拟蟹守螺、菲律宾蛤仔、长腕和尚蟹、纵带滩栖螺、青蛤、南海鸭嘴蛤、文蛤和红树蚬等8种, 年均密度和生物量分别为203个×m-2和276.58g×m-2, 其中软体动物占69.58%和83.73%。盐度和底质类型因素是影响群落密度、生物量和群落指数的主要环境因子, 季节和潮带因素影响较小。其中盐度对密度和生物量的影响大于底质类型, 但底质类型对种类多样性指数、丰富度指数、均匀度指数和物种丰度等群落指数的影响大于盐度。聚类分析和多维尺度分析可以将群落划分为3个类型, 基本对应于盐度和底质类型影响。从多样性水平判断防城河口湾潮间带总体上处于中度扰动之中, 盐度、底质类型和人为干扰等因素综合影响了该湾潮间带动物群落的空间分布格局。
防城河口湾; 潮间带大型底栖动物; 群落结构; 环境扰动
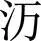
1 材料与方法
1.1 调查站位设置与采样
在防城河口湾潮间带滩涂共设置7条调查断面(对应编号S1至S7, 断面示意见图1), 每条断面在高、低潮带各设1~2个站位, 中潮带布设3个站位。各潮带的生境因子见表1, 21处采样潮带的底质类型有8处为沙泥质裸滩, 5处为红树林滩涂, 5处为沙质裸滩, 3处为淤泥质裸滩。分别于2016年5月、8月、11月和2017年2月的当地大潮期, 开展潮间带大型底栖动物定量定性采样。在每个站位采集4个定量样方, 样方大小为25cm×25cm, 采样深度30cm。挖取样方框范围内的沉积物放入二层网目均为1mm的套筛内反复冲洗, 拣出滞留网上肉眼可见动物, 并将残渣带回实验室在显微镜下挑出所有动物。所得样品用5%甲醛固定5d后进行种类鉴定、计数和称重(精确度为0.001g)。每条断面的站位密度和生物量按其所在的潮带归类, 取潮带平均值, 作为各潮带的栖息密度(个×m-2)和生物量(g×m-2)。定性取样在等高线内进行带状取样。野外调查时使用YSI 650多参数水质仪测定退潮后沉积物渗出水及涨潮时上覆水的pH和盐度, 结果见表1。
1.2 数据处理
1.2.1 群落多样性和相似性指数计算
群落多样性指数和相似性指数的算式如下:
Shannon-Weaver多样性指数(′):
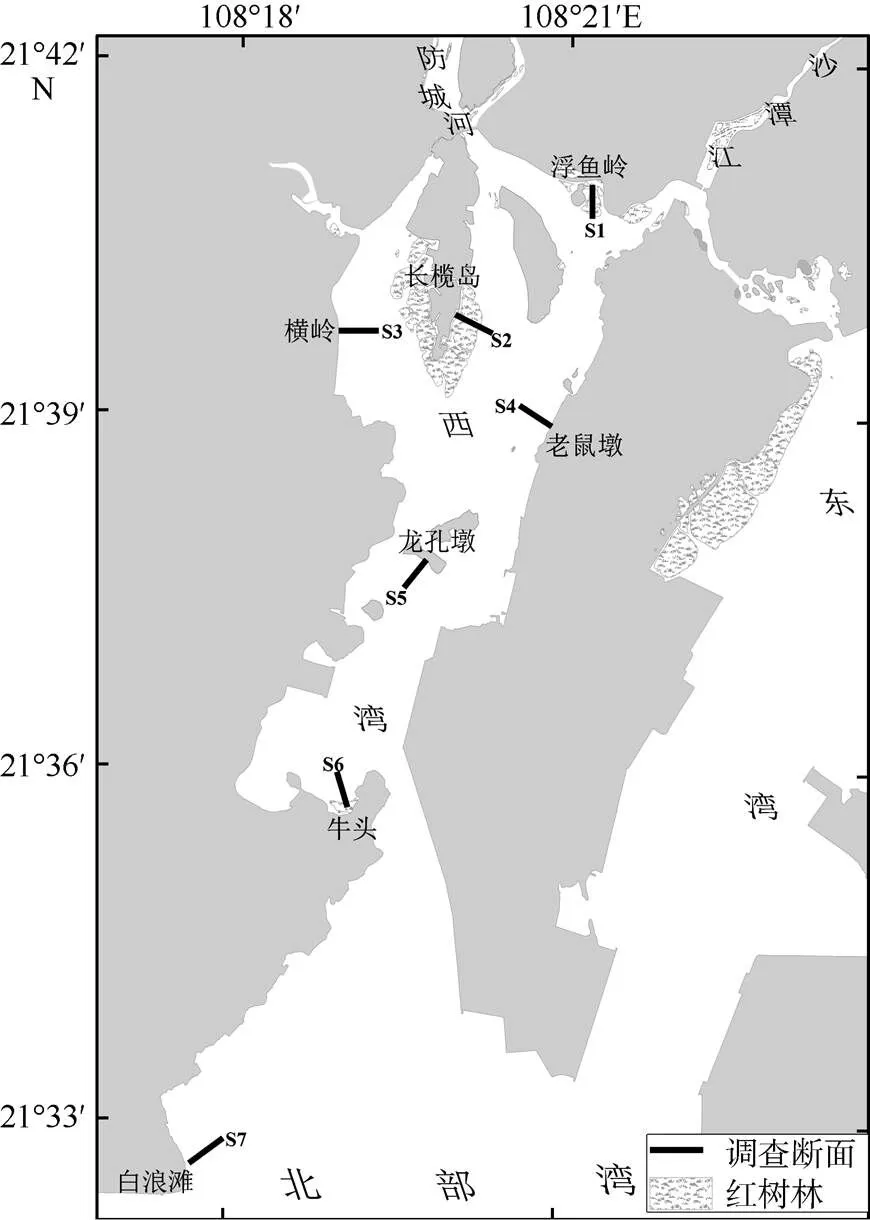
图1 防城河口湾潮间带大型底栖动物调查断面示意图
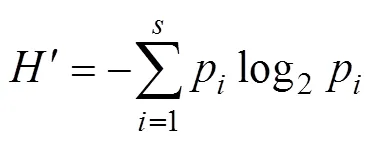
Margalef丰富度指数():
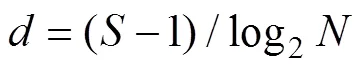
Pielou均匀度指数():
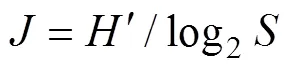
式中:为某调查潮带的所有物种的个体数之和;为该潮带定量样方鉴定出的物种数目;p为某潮带中第个种的个体数占该潮带总个数的比例。
物种相似性指数S(单位: %):
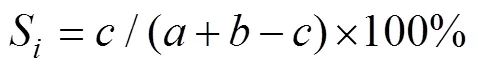
式中:、分别为某两个断面的物种数;为两个断面共有的物种数。
1.2.2 群落相对重要值计算
相对重要值(单位: %)计算采用如下算式:

式中:n为第种密度(单位: 个×m-2);w为第种生物量(单位: g×m-2)。设定≥1%的种群为优势种群。
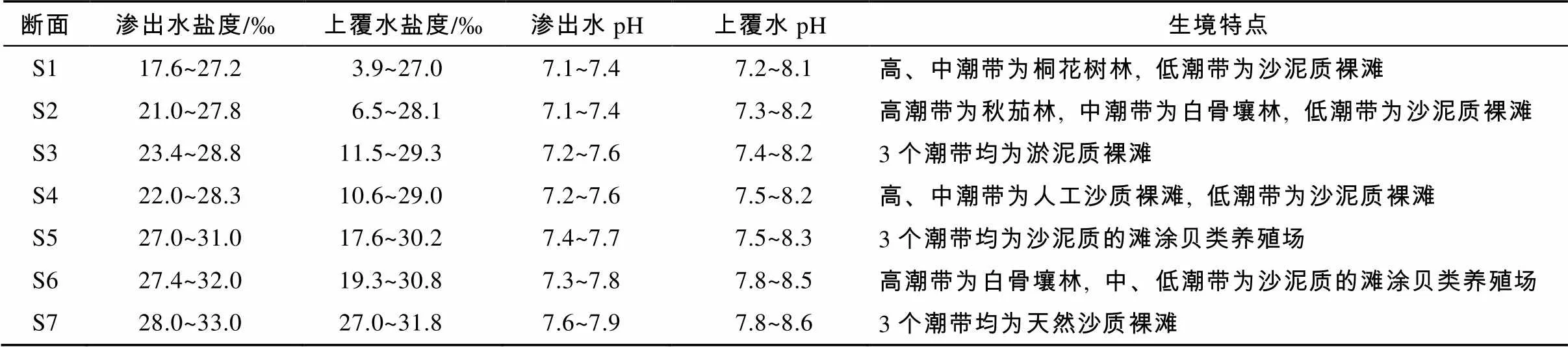
表1 防城河口湾潮间带大型底栖动物调查区域的盐度、pH和生境类型
1.2.3 数据分析
采用IBM SPSS Statistics 23.0软件对不同生境潮间带大型底栖动物群落的密度、生物量和群落指数进行方差分析, 将4个季度各21处潮带的密度数据作4次方根转换, 进行系统聚类分析和非线性多维尺度分析。
2 结果与分析
2.1 群落种类组成
4个季度共采集到潮间带大型底栖动物10门14纲252种, 以软体动物门、节肢动物门和环节动物门为主。其中, 软体动物门有2纲124种, 占总种数的49.2%, 双壳纲和腹足纲分别有82种和42种。节肢动物门有2纲65种, 占25.8%, 甲壳纲和肢口纲分别有63种和2种。环节动物门有1纲40种, 占15.9%, 均属多毛纲。脊索动物门有1纲14种, 占5.6%, 均属硬骨鱼纲。其他6门有9种, 包括棘皮动物门3种(含海胆纲1种、海参纲2种), 星虫动物门2种(含革囊星虫纲和方格星虫纲各1种), 及扁形动物门涡虫纲、纽形动物门无针纲、尾索动物门有尾纲和腕足动物门无关节纲各1种。
各断面采集到的动物种数差异较大, S1获100种, S2获82种, S3获64种, S4获48种, S5获103种, S6获82种, S7获108种。位于湾口的S7种数最多, 位于湾中上部的S4最少。断面S7与其他断面之间的相似度低, 尤其是S7与S3之间相似性指数仅10.3%。S7位于湾口为沙质滩涂, S3位于湾上部为淤泥质滩涂, 底质类型和盐度差异均较大。S1与S2之间的相似性指数最大, 达42.2%, S5与S6之间次之, 为35.0%, 空间相邻且底质类型近似的断面间相似程度较大。
虽然各断面种数差异较大, 但各类群种数占比的大小顺序却一致, 均表现为: 软体动物门>节肢动物门>环节动物门>其他门类。软体动物门、节肢动物门和环节动物门是防城河口湾潮间带动物群落的优势类群。自湾顶至湾口, 主要优势种群呈现出由适应河口低盐种群向适应湾口高盐种群逐渐过渡的趋势: 在湾顶, 以斜肋齿蜷、中国绿螂、弓绿螂和红树蚬等河口种占优势, 在外湾则多见古明志圆蛤、四角蛤蜊、突畸心蛤、高蛤蜊和紫藤斧蛤等适应高盐种群。
2.2 群落优势种
全年防城河口湾7个断面潮间带动物群落中相对重要值≥1%的优势种有44种, 包括软体类28种, 甲壳类10种, 多毛类和其他类群各3种(表2)。软体类在5个断面、甲壳类在2个断面为最大优势类群, 软体类的优势地位显著。除了S3和S7外, 其他5个断面最大优势种的相对重要值均超过50%, 多样化程度相对较弱。7个断面中, S1、S2、S6的最大优势种均为珠带拟蟹守螺, S3的是莱氏异额蟹, S4的是长腕和尚蟹, S5的是菲律宾蛤仔, S7的是古明志圆蛤。珠带拟蟹守螺在红树林和沙泥质滩涂生境优势突出, 在S1、S2和S6的相对重要值分别达到54.86%、77.95%和66.17%, 但在盐度较高且沙质的湾口S7则几近消失。全年统计整个调查区域的8个优势种及其重要值分别为: 珠带拟蟹守螺(52.28%)、菲律宾蛤仔(19.15%)、长腕和尚蟹(7.29%)、纵带滩栖螺(6.89%)、青蛤(3.74%)、南海鸭嘴蛤(2.61%)、文蛤(2.44%)和红树蚬(1.04%)。
2.3 群落密度与生物量
防城河口湾潮间带的动物群落年平均密度为203个×m-2, 其中软体类占69.58%, 甲壳类占22.38%, 多毛类占5.36%, 其他类群占2.68%。空间分布呈现河口湾中部(S5、S6)较高、南北较低的格局(表2), 密度变化较大, 方差分析表明断面之间差异极显著(6, 77=7.626,<0.01)。4类底质类型之间的密度差异极显著(3, 80=7.572,<0.01), 分别为: 红树林滩涂(270个×m-2)、沙泥质裸滩(266个×m-2)、沙质裸滩(125个×m-2)、淤泥质裸滩(44个×m-2)。4个季节之间差异不显著(3, 80=0.028,>0.05), 高、中、低潮带之间密度差异也不显著(2, 81=0.928,>0.05)。盐度与底质类型对密度分布的影响程度相近。盐度、潮带、底质类型和季节4类因素之间, 盐度与底质类型、盐度与潮带有交互作用, 其他因素间均无交互作用。
调查区域的年均生物量为276.58g×m-2, 软体类、甲壳类、其他类群和多毛类分别占83.73%、11.36%、4.36%和0.55%。各断面生物量分布规律与密度大致相似(表2), 断面之间生物量差异极显著(6, 77=6.346,<0.01)。底质类型之间生物量差异显著(3, 80=3.936,<0.05), 沙泥质裸滩(411.69g×m-2)>红树林滩涂(261.71g×m-2)>淤泥质裸滩(155.63g×m-2)>沙质裸滩(147.83g×m-2)。季节之间、潮带之间的生物量差异均不显著(3, 80=1.048,2, 81= 1.560,>0.05)。盐度对生物量分布的影响程度大于底质类型。4类因素间均无交叉作用。
2.4 群落指数
全年定量采集获得的各潮带平均物种丰度为8.2种; 8~11种者较多, 占41.7%; 16种以上者占3.6% (图2)。底质类型之间的物种丰度差异极显著(3, 80=22.098,<0.01); 断面之间物种丰度差异极显著(6, 77=15.430,<0.01);潮带之间、季节之间物种丰度差异均不显著(2, 81=3.060,3, 80=0.796,>0.05)。底质类型对物种丰度分布的影响程度大于盐度。底质类型与潮带、盐度与潮带有交互作用, 其他因素均无交叉作用。
多样性指数′大于3者仅占总数8.3%, 其中S5有3例, S1至S4各有1例(图2);′值在2~3者占了50.0%。底质类型之间、断面之间′值的差异极显著(3, 80=18.716,6, 77=8.195,<0.001), 底质类型的影响程度大于盐度。潮带之间、季节之间的′值差异均不显著(2, 81=1.452,3, 80=0.287,>0.05)。底质类型与潮带、盐度与潮带有交互作用, 其他因素均无交叉作用。
丰富度指数值分布较集中在1~2的区间, 占61.9% (图2)。底质类型之间、断面之间值差异均极显著(3, 80=21.296,6, 77=10.654,<0.01)。潮带之间值差异显著(2, 81=3.225,<0.05)。季节之间值差异不显著(3, 80=0.541,>0.05)。对值分布影响程度的大小顺序依次为底质类型、盐度和潮带。底质类型与潮带、盐度与潮带有交互作用, 其他因素均无交叉作用。
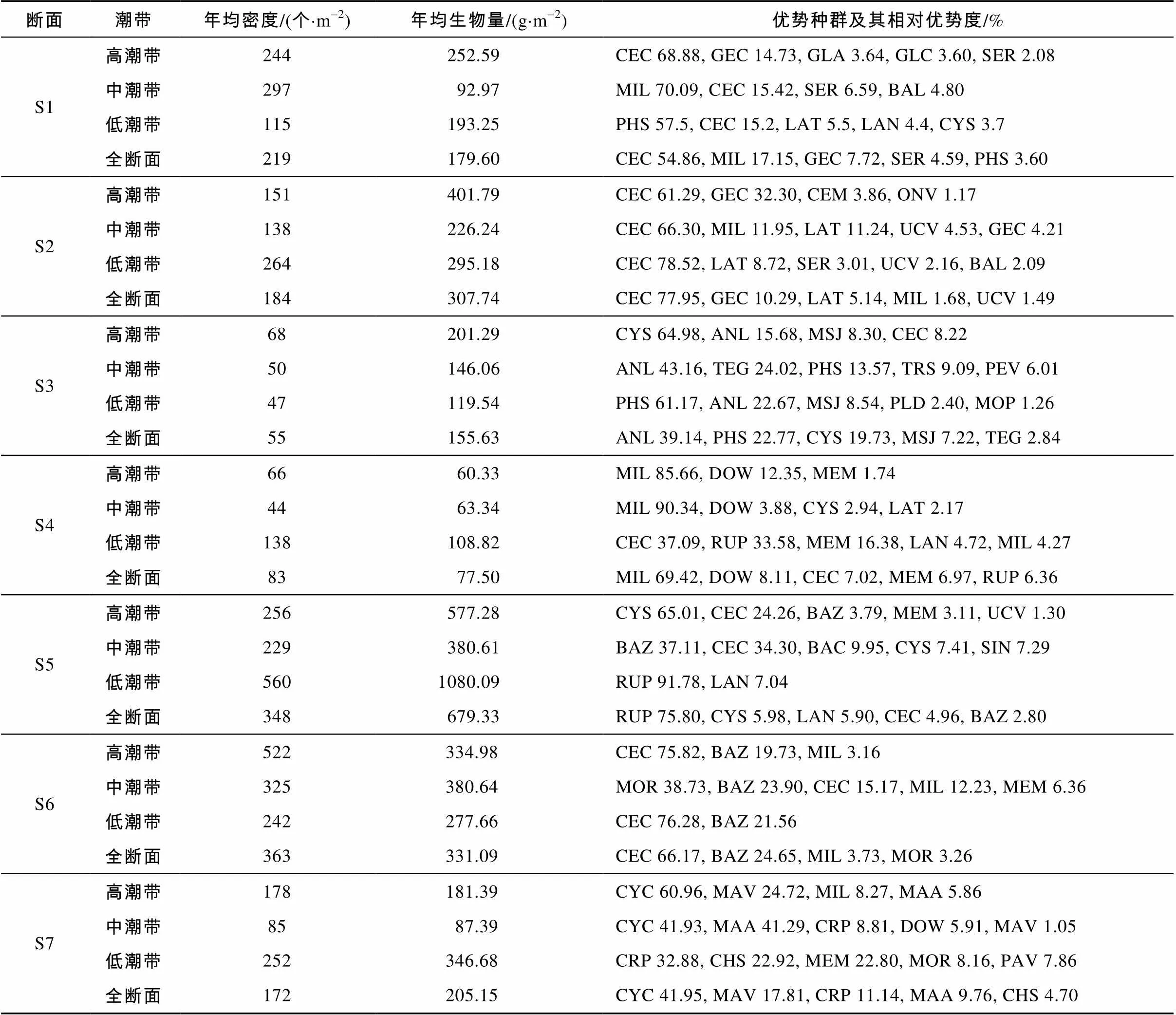
表2 防城河口湾潮间带大型底栖动物群落密度、生物量及优势种群相对重要值
注: ANL=莱氏异额蟹, BAC=古氏滩栖螺, BAL=潮间藤壶, BAZ=纵带滩栖螺, CEC=珠带拟蟹守螺, CEM=小翼拟蟹守螺, CHS =紫藤斧蛤, CRP=突畸心蛤, CYC=古明志圆蛤, CYS =青蛤, DIP=拟脊活额寄居蟹, DOW=韦氏毛带蟹, FEP=长毛明对虾, GEC=红树蚬, GLA=弓绿螂, GLA=白色吻沙蚕, GLC=中国绿螂, ILN=宁波泥蟹, LAN=南海鸭嘴蛤, LAT=截形鸭嘴蛤, LIA=亚氏海豆芽, MAA=高蛤蜊, MAV=四角蛤蜊, MEL=长足长方蟹, MEM=文蛤, MER=斧文蛤, MIL=长腕和尚蟹, MOP=菲律宾偏顶蛤, MOR=红明樱蛤, MSJ=日本大眼蟹, NIJ=幼形亮樱蛤, NOL=背蚓虫, ONV=石磺, PAV=南海毛满月蛤, PEV=翡翠贻贝, PHS=针骨沙鸡子, PLD=杜氏阔沙蚕, RUP=菲律宾蛤仔, SER=斜肋齿蜷, SIN=裸体方格星虫, TEG=泥蚶, TRS=亚光棱蛤, UCA=弧边招潮, UCV=凹指招潮
均匀度指数值多出现在0.75~1.00区间, 占54.8% (图2)。底质类型之间、断面之间值差异均显著(3, 80=4.601,6, 77=3.991,<0.05), 底质类型的影响程度大于盐度。潮带之间、季节之间值差异均不显著(2, 81=0.095,3, 80=0.028,>0.05)。因素间均无交叉作用。
部分群落指数之间存在密切关系,与物种丰度、与′的相关系数分别达0.887和0.834,′与物种丰度、′与的相关系数分别达0.644和0.610, 均极显著(<0.01,=84)。
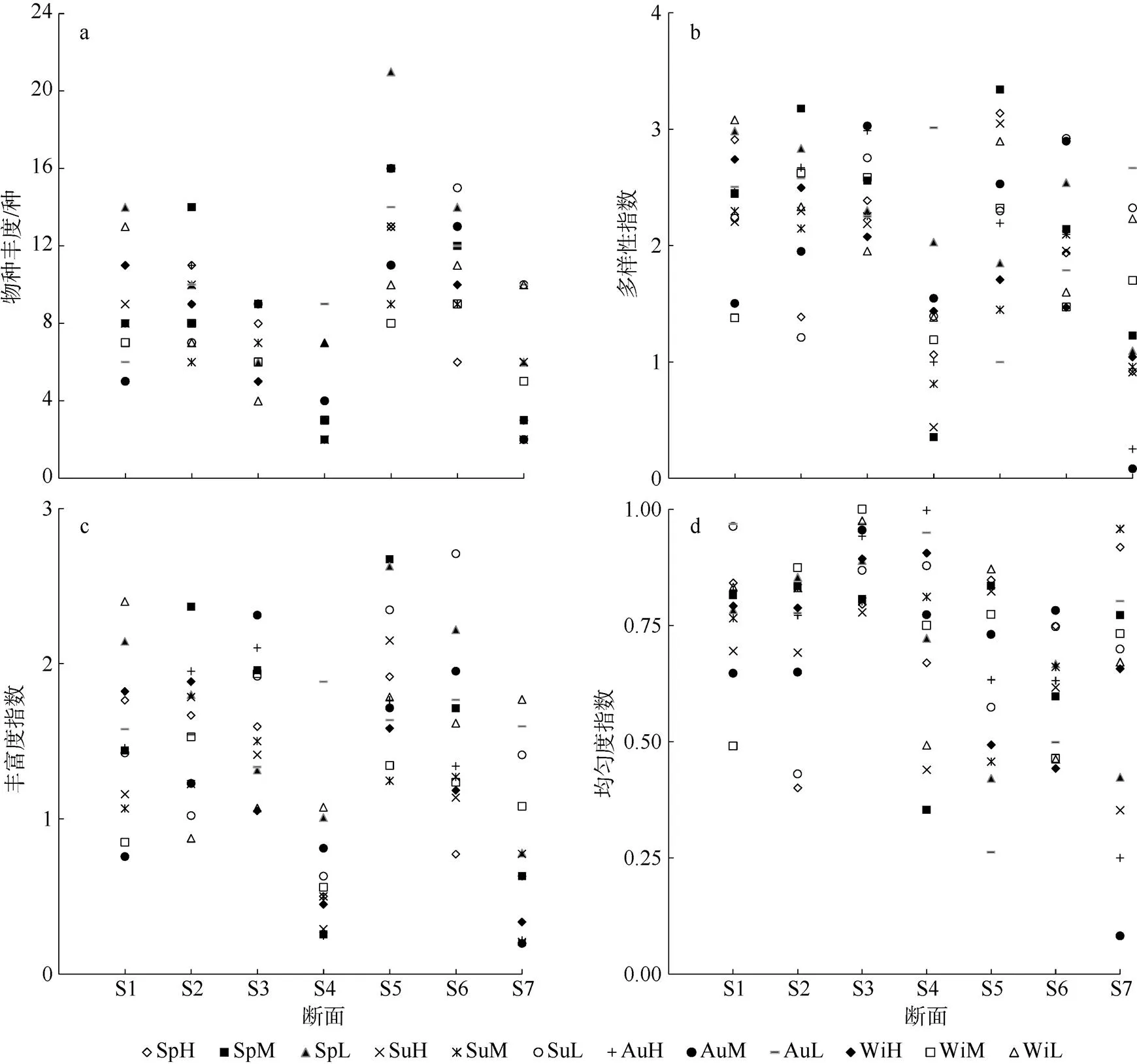
图2 防城河口湾全年各潮带大型底栖动物群落物种丰度(a)、种类多样性指数(b)、丰富度指数(c)、均匀度(d)
图例3个字母中前2个为季节的英文缩写, Sp春季, Su夏季, Au秋季, Wi冬季; 后1个为所在潮带英文缩写, H高潮带, M中潮带, L低潮带
Fig. 2 Indexes of species abundance (a), species diversity′ (b), Richness(c), and Evenness(d) of macrozoobenthic communities seasonally sampled in the intertidal zone of Fangchenghe Estuary. Among the three letters in each legend, the first two letters are the abbreviation of sampling season: Sp = Spring, Su = Summer, Au = Autumn, and Wi = Winter; and the last letter is the abbreviation of tidal zone: H = high tidal zone, M = middle tidal zone and L = low tidal zone
2.5 聚类分析与多维尺度分析
聚类图和排序图见图3。聚类图显示按欧氏距离10%可划分为3个群: 第一个群主要包括S3和S4的潮带, 生境为淤泥质滩涂和沙质滩涂类型, 匍匐生活型、底游生活型、凿穴生活型动物种类和数量很低甚至为0, 各潮带年均密度明显较低; 第二个群是河口上游以珠带拟蟹守螺为优势种的群, 密度稍高于第一个群; 第三个群以S5和S6的潮带为主, 生境为异质性程度较高的沙泥质滩涂及红树林, 该群所包含的潮带最多, 年均密度较高。此外还有3个潮带呈较独立星散分布, 包括菲律宾蛤仔(S5L)、珠带拟蟹守螺(S6H)大量繁衍的潮带, 及季节分布格局迥异于其他潮带的S7H。
从MDS排序图中不同程度分离的潮带分布可看出, MDS排序结果与群落聚类分析结果基本一致, 进一步印证了聚类分析的结果。
3 讨论
3.1 防城河口潮间带动物群落结构特征
防城河口湾潮间带大型底栖动物群落种类组成以软体动物门、节肢动物门和环节动物门为主体, 这与已报道的广西其他海域类似(何斌源等, 1998, 2004, 2013; 何祥英等, 2012; 许铭本等, 2015)。但本文调查获得10门252种, 远多于广西其他海区的报道, 例如何斌源等(1998)在珍珠湾红树林区设5站位调查获84种, 何斌源等(2004)在钦州湾潮间带设4断面11站位调查获70种, 何祥英等(2012)在北仑河口红树林区设4断面15站位调查获106种, 许铭本等(2015)在竹山红树林区设3断面10站位调查获63种。比较定量样方获得种数, 本文设7断面40站位调查获157种, 与何斌源等(2013)在廉州湾潮间带设12断面33站位调查获156种接近。可见, 生境多样性、取样站位数、地理位置甚至采样方法等因素, 均可影响我们的区域生物多样性调查结果。
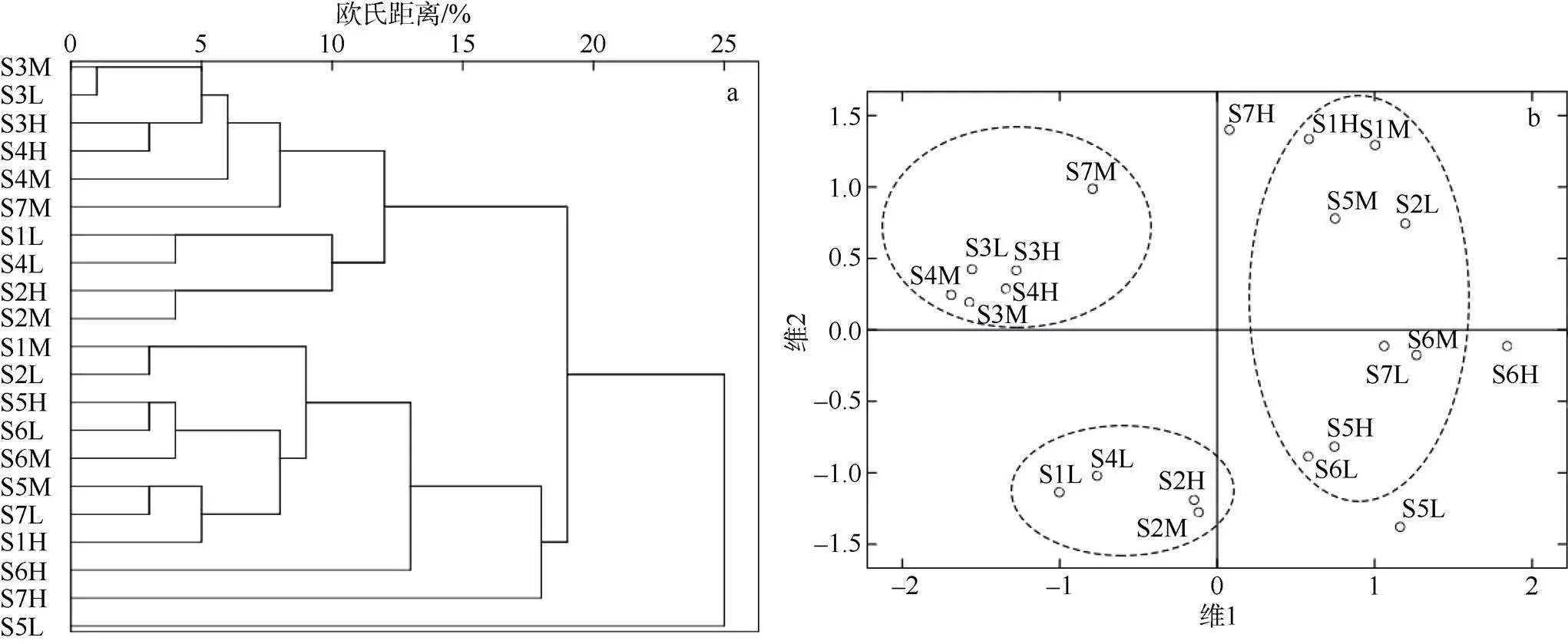
图3 防城河口湾潮间带大型底栖动物群落系统聚类(a)及MDS分析图(b)
H为高潮带; M为中潮带; L为低潮带; 图b中虚线圈显示MDS分析划分的3个主要的群
Fig. 3 The hierarchical cluster dendrogram (a) and 2-dimentional MDS ordinal configuration (b) of macrozoobenthic communities in the intertidal zone of Fangchenghe Estuary.H= high tidal zone, M= middle tidal zone and L= low tidal zone, Three dotted circles in figure b represent three major groups based on the MDS ordinal
防城河口湾潮间带底栖动物群落中很多为广布种, 部分为河口低盐种。在湾顶常见斜肋齿蜷、中国绿螂、弓绿螂、红树蚬、河蚬、宁波泥蟹、伍氏拟厚蟹等河口低盐种, 且有时形成较大优势; 在湾口则多见古明志圆蛤、四角蛤蜊、突畸心蛤、高蛤蜊和紫藤斧蛤等适应较高盐度的动物。湾顶断面S1与湾口断面S7群落之间共有种仅34种, 均为广布种, 相似性指数仅19.5%, 可见两地动物群落组成结构差异非常大。同时一些种类在河口湾广泛地分布, 如珠带拟蟹守螺、长腕和尚蟹、韦氏毛带蟹等。群落组成在盐度适应生态类型上的变化是河口湾的一种生物特征, 袁兴中等(2002)在长江口潮滩湿地调查获得大型底栖动物68种, 河口低盐种、半咸水种和淡水种共存; 何斌源等(2013)发现随着水体盐度上升, 南流江入海口廉州湾红树林和裸滩的潮间带动物群落优势种出现了由适应低盐环境向适应高盐环境的种类组成变化。
河口多变的环境因子仅适合为数不多的种群大量繁衍, 通常生物群落主要优势种突出。防城河口湾潮间带动物群落中相对重要值≥1%的优势种有珠带拟蟹守螺、菲律宾蛤仔等8种, 仅占调查区域总种数252种的3.2%。通常珠带拟蟹守螺在红树林及其邻近滩涂上优势度非常显著, 该种群聚性强,具有很高的频度、密度和生物量, 重要值达52.28%, 超过其他所有种类之和。重要值次之的菲律宾蛤仔则非常集聚在断面S5低潮带, 春季达856个×m-2和1841.48g×m-2, 分别占该种全年总量的53.4%和86.3%。在其他海区潮间带也常见优势种集聚于少数种群的现象, 例如, 在广西廉州湾潮间带上珠带拟蟹守螺、弧边招潮等少数物的种优势度非常显著(何斌源等, 2013); 辽宁东营市河口区潮间带动物群落中泥螺和文蛤异常高于其他种群(顾炎斌等, 2013); 阿根廷Bahía Blanca河口沙滩潮间带动物群落中, 豆蟹科的优势度达64.09%, 其他任一种均不超过10% (Carcedo et al, 2015); 福建坛南湾潮间带上肋昌螺有着很高的密度, 某站高达3600个×m-2(李国强等, 2017)。优势种集聚于少数种群往往导致群落多样性指数′较低, 防城河口湾潮间带动物群落多样性指数′大于3的潮带数仅占总数8.3%, 绝大部分处在1~3的范围内, 群落结构受到中度扰动。
3.2 防城河口潮间带动物群落结构的影响因素与管理控制
生境相对稳定是维持生物群落结构的关键因素, 然而河口湾是海陆作用最为剧烈、生态结构最为复杂的区域, 同时也是人为干扰最频繁、污染最严重的区域, 而且人为因素影响呈现超过自然因素的趋势, 引起生态质量及生态服务功能逐步衰退。已有研究表明, 潮间带底栖动物群落的组成性质、区域分布及数量特征等与其盐度、底质类型、季节、潮带和相关生物等自然因素(蔡立哲等, 2001; 袁兴中等, 2002; Edgar et al, 2002; Chapman et al, 2004; Occhipinti-Ambrogi et al, 2005; Fujii, 2007; Otani et al, 2008; Picanço et al, 2014; Carcedo et al, 2015), 以及疏浚、围填海、排污等人为因素(Belan, 2003; 叶属峰等, 2004; 金亮等, 2007; 周进等, 2012; 张莹等, 2012; 贺心然等, 2015; 崔磊等, 2017)密切相关。
通常多种自然和人为因素共同影响潮间带底栖动物群落种类组成和群落数量指标的空间差异, 同时诸多因素之中有主导因素和次要因素的区别。本文结果表明: 盐度、底质类型、季节和潮带等4种自然因素对防城河口湾潮间带动物群落数量指标造成的影响不一。季节和潮带因素对各个数量指标造成的影响较小, 而盐度和底质类型因素则影响显著至极显著, 后两个因素综合主导了潮间带动物的物种分布格局及群落结构, 盐度对密度和生物量分布的影响尤其突出。由湾顶至湾口, 防城河口湾水体盐度由较低且激烈变动逐渐升高且稳定(表1)。盐度差异大是河口区显著的宏观特征, 可观察到盐度主导河口湾生物群落中的河口种、海洋种和淡水种出现与否及数量多寡。袁兴中等(2002)指出, 大尺度上长江口潮滩湿地底栖动物物种及功能群分布格局主要决定于盐度梯度。厉红梅等(2003)研究发现, 靠近深圳河口的潮间带底栖动物群落与红树林区附近潮间带底栖动物群落结构有明显的差异, 群落结构组成随盐度梯度而呈现连续的变化。何斌源等(2013)报道, 广西南流江口裸滩上底栖动物群落优势种随水体盐度升高而变化, 由穴居生活型蟹类(弧边招潮)占优转向底上生活型软体类(珠带拟蟹守螺)等, 且断面种数与盐度具有正相关关系。
底质类型对防城河口湾潮间带动物群落的种类多样性指数′、丰富度指数均匀度和潮带物种丰度等4个群落指数的影响超过了盐度。本文物种丰度并没有出现随盐度升高而增多的现象(图2), 这与长江口(袁兴中等, 2002)、南流江口(何斌源等, 2013)、珠江口(张敬怀, 2014)等河口水域不一致。该海湾的物种丰度低值区出现在调查区域中上部的断面S3和S4, 其上、下游区域均出现高值区, 密度和生物量也出现类似的分布格局, 这主要由底质类型因素引起。由于防城河口湾的西航道被养殖堤围阻塞, 长榄岛至横岭一带滩涂(图1中的S3断面及周边)水交换不畅, 加速外源细颗粒物质沉积形成了淤泥质滩涂; 断面S4位于河口湾东岸的人造沙滩, 堆积在高潮带至潮上带的沙粒受冲刷不断向下流失, 造成生境不稳定。聚类分析与MDS分析均表明: 断面S3和S4的潮间带动物群落结构趋向于相似, 尽管它们的底质类型差异很大, 但均为不稳定生境, 容易导致某些功能群数量锐减甚至缺失, 如: 断面S3和S4的匍匐生活型动物(腹足类)生物量占比仅分别为2.4%和6.4%, 而其他断面占比在11.1%~46.2%之间; 底游生活型、凿穴生活型动物数量极少甚至为0。在深圳湾淤泥质滩涂一般多毛类比重显著上升, 密度很高, 但生物量极低(蔡立哲等, 2001; 厉红梅等, 2003); 表明底质类型因素作用非常显著。
通过比较和统计分析可了解多个生态因素的综合影响作用。厉红梅等(2004)通过4个季度采样得出结论, 季节和潮带是影响深圳湾底栖动物群落结构组成时空变异的主要因素, 同时也与污染程度差异和距河口的远近等次要因素有关。赵永强等(2009)分析椒江口滩涂大型底栖动物群落格局与多样性, 认为在河口区自然环境因素中盐度、底质结构等变化是影响生物分布的主要环境因子。侯森林等(2011)研究发现, 潮带和湿地植被是影响射阳河口潮间带大型底栖动物群落结构组成的两个重要因素。Picanço等(2014)报道沉积物的细沙组分和有机质含量显著影响葡萄牙Minho河口盐沼潮间带动物群落结构, 而重金属含量的影响不显著。Carcedo等(2015)分析表明阿根廷Bahía Blanca河口沙滩的盐度、温度、波浪周期、波高、粒度等指标与不同的群落指数有显著相关性。李晓静等(2016)认为水温、盐度和总磷含量等环境因子与山东烟台大沽夹河口及邻近海域底栖动物群落的丰度和生物量空间分布特征相关性较大。当然, 本文认为还需开展更深入的工作解决如何分辨多种影响因子的程度与权重问题。
人为扰动引起生境的理化因素和生物因素变化而对底栖生物群落产生影响。叶属峰等(2004)调查发现, 由于长江口经常性疏浚, 该水域底栖生物种数由1978年的153种减少至2002年的19种, 密度和生物量也急剧降低, 引起底栖生态系统结构与功能的改变, 而且通过食物链关系, 导致长江口渔业资源全面衰退。Rehitha等(2017)研究表明维护性疏浚导致印度Cochin河口疏浚区底栖动物群落密度、生物量和多样性指数显著地低, 且属颤蚓异常丰富, 而双壳类和端足类很少。在污染严重的深圳湾北岸红树林外泥滩上多毛类和寡毛类等耐高有机负荷的种类最多, 年均生物量仅有18.52g×m-2, 但年均密度达到103~104个×m-2水平(金亮等, 2007)。在福建东北部的三都澳海湾养殖使得污染逐年加重, 呈现富营养化趋势, 潮间带底栖动物年均密度较高, 可达到121.52个×m-2, 但年均生物量仅有7.00g×m-2, 与自然水域差别明显(周进等, 2012)。张莹等(2012)报道小清河口底栖动物群落的多样性指数′与铵盐、硝酸盐、铜和汞含量呈负相关, 认为生活污水和工业废水的排放对河口区底栖动物群落产生了负面影响。贺心然等(2015)研究表明江苏灌河口潮间带底栖动物种数、密度、′等都比邻近海域的低, 认为源自灌河沿岸化工排放的污染物如敌敌畏、有机氯农药、半挥发性有机物、镉、汞、锌和总铬等造成了显著或较大影响。崔磊等(2017)连续调查发现围填海工程不但导致广东淇澳岛附近水域重金属污染及富营养化, 而且造成生物多样性降低及优势种和群落结构的变化, 影响群落稳定性。上述研究所在区域生境条件及关注的环境指标不一样, 其揭示的影响潮间带动物群落的胁迫机制各具区域特殊性, 当然同时也存在普遍性。
如果我们采取有效措施将人为扰动控制在一定程度, 河口湾生物有可能逐渐适应变化后的环境而建立起一个新的生态系统。蔡立哲等(2007)报道在2000年初, 深圳市在落马洲河段两端与深圳河连接处建造了水闸, 截断了污染物进入落马洲河段的通道, 截污后两年监测发现该河段大型底栖动物从无到有, 种数、密度和生物量从少到多, 群落结构处于向多样性恢复的过程中。刘修泽等(2011)发现辽宁旅顺南部的基岩海岸潮间带大型底栖动物群落平均生物量达4312.5g×m-2, 平均栖息密度达2966个×m-2, 原因是该区域为确权海域, 生物资源管理较科学合理,人为干预较少, 采捕经济种时执行取大留小。本文中受到管理的滩涂贝类养殖区断面S5和S6, 采样潮带平均种数分别达12.8种和10.8种, 多样性指数也显著高于其他断面, 表明管理措施促进底栖动物群落的维持和发展。在国家海洋局支持下, 目前有关部门正准备在防城河口湾实施“蓝色海湾”整治行动, 水道疏浚、拆除原有堤围、生态化海堤建设等工程将急剧改变河口湾水动力、水质和沉积物状况, 必须对其生态影响进行严密跟踪调查监测, 及时评估工程效果, 编制和实施海域生态保护及修复计划。
蔡立哲, 厉红梅, 林鹏, 等, 2001. 深圳河口潮间带泥滩多毛类的数量变化及环境影响[J]. 厦门大学学报(自然科学版), 40(3): 741–750. CAI LIZHE, LI HONGMEI, LIN PENG, et al, 2001. Analysis of environmental effect and polychaete quantitative variations on intertidal mudflat in Shenzhen estuary[J]. Journal of Xiamen University (Natural Science), 40(3): 741–750 (in Chinese with English abstract).
蔡立哲, 林鹏, 厉红梅, 2007. 截污后深圳河落马洲段大型底栖动物群落的恢复过程[J]. 应用与环境生物学报, 13(4): 497–500. CAI LIZHE, LIN PENG, LI HONGMEI, 2007. Restoration process of macrofaunal community at Luomazhou section of the Shenzhen River after pollution interception[J]. Chinese Journal of Applied & Environmental Biology, 13(4): 497–500 (in Chinese with English abstract).
崔磊, 吕颂辉, 董悦镭, 等, 2017. 围填海工程对淇澳岛附近水域环境因子与生物群落的影响[J]. 热带海洋学报, 36(2): 96–105. CUI LEI, LÜ SONGHUI, DONG YUELEI, et al, 2017. Influence on the biological community and environmental factors around Qi’ao Island caused by reclamation project[J]. Journal of Tropical Oceanography, 36(2): 96–105 (in Chinese with English abstract).
顾炎斌, 宋文豪, 姚韡远, 等, 2013. 东营市河口区潮间带大型底栖动物群落结构特征[J]. 海洋环境科学, 32(5): 702–706. GU YANBIN, SONG WENHAO, YAO WEIYUAN, et al, 2013. Intertidal macrobenthic community structural features in Hekou district[J]. Marine Environmental Science, 32(5): 702–706 (in Chinese with English abstract).
何斌源, 范航清, 张振日, 1998. 珍珠港红树林大型底栖动物生态的初步研究[M]//何其锐. 南海资源开发研究. 广州: 广东经济出版社: 1036–1048. HE BINYUAN, FAN HANGQING, ZHANG ZHENRI, 1998. Preliminary study on the ecology of mangrove macrobenthos in Pearl bay, Guangxi [M]//HE QIRUI. Resources development and research in South China Sea. Guangzhou: Guangdong Economy Press: 1036–1048 (in Chinese).
何斌源, 邓朝亮, 罗砚, 2004. 环境扰动对钦州港潮间带大型底栖动物群落的影响[J]. 广西科学, 11(2): 143–147. HE BINYUAN, DENG CHAOLIANG, LUO YAN, 2004. Effect of environmental fluctuation on macrobenthos community in the intertidal flats of Qinzhou Harbor[J]. Guangxi Sciences, 11(2): 143–147 (in Chinese with English abstract).
何斌源, 赖廷和, 王欣, 等, 2013. 廉州湾滨海湿地潮间带大型底栖动物群落次级生产力[J]. 生态学杂志, 32(8): 2104–2112. HE BINYUAN, LAI TINGHE, WANG XIN, et al, 2013. Secondary productivity of benthic macrofaunal community in intertidal zone of Lianzhou Bay, China[J]. Chinese Journal of Ecology, 32(8): 2104–2112 (in Chinese with English abstract).
何祥英, 苏搏, 许廷波, 等, 2012. 广西北仑河口红树林湿地大型底栖动物多样性的初步研究[J]. 湿地科学与管理, 8(2): 44–48. HE XIANGYING, SU BO, XU TINGBO, et al, 2012. Macrobenthic biodiversity in mangrove wetland at the estuary of Beilun River in Guangxi. Wetland Science & Management, 8(2): 44–48 (in Chinese with English abstract).
贺心然, 陈斌林, 高文婕, 等, 2015. 灌河口潮间带及其入海河段秋季大型底栖动物生态学研究[J]. 海洋科学, 39(5): 28–35. HE XINRAN, CHEN BINLIN, GAO WENJIE, et al, 2015. Ecological studies of macrobenthos in the intertidal zone and near sea section of Guan River in autumn[J]. Marine Sciences, 39(5): 28–35 (in Chinese with English abstract).
侯森林, 余晓韵, 鲁长虎, 2011. 盐城自然保护区射阳河口潮间带大型底栖动物空间分布与季节变化[J]. 生态学杂志, 30(2): 297–303. HOU SENLIN, YU XIAOYUN, LU CHANGHU, 2001. Spatial distribution and seasonal variation of macrobenthos in intertidal flat of Sheyang estuary, Yancheng Nature Reserve[J]. Chinese Journal of Ecology, 30(2): 297–303 (in Chinese with English abstract).
金亮, 蔡立哲, 周细平, 等, 2007. 深圳湾北岸泥滩大型底栖动物次级生产力研究[J]. 台湾海峡, 26(3): 415–421. JIN LIANG, CAI LIZHE, ZHOU XIPING, et al, 2007. Secondary production of macrobenthos on the mudflat of northern Shenzhen Bay[J]. Journal of Oceanography in Taiwan Strait, 26(3): 415–421 (in Chinese with English abstract).
李国强, 叶伟鹏, 余怀勇, 等, 2017. 平潭岛中国鲎保护区沙质潮间带的大型底栖动物群落[J]. 海洋环境科学, 36(2): 179–185. LI GUOQIANG, YE WEIPENG, YU HUAIYONG, et al, 2017. Community of benthic macrofauna on sandy intertidal zone in Chinese horseshoe crab reserve in Pingtan Island, China[J]. Marine Environmental Science, 36(2): 179–185 (in Chinese with English abstract).
厉红梅, 李适宇, 蔡立哲, 2003. 深圳湾潮间带底栖动物群落与环境因子的关系[J]. 中山大学学报(自然科学版), 42(5): 93–96. LI HONGMEI, LI SHIYU, CAI LIZHE, 2003. Relationship between benthic community and environmental factors in Shenzhen Bay[J]. Acta Scientiarum Naturalium UniversitatisSunyatseni, 42(5): 93–96 (in Chinese with English abstract).
厉红梅, 孟海涛, 2004. 深圳湾底栖动物群落结构时空变化环境影响因素分析[J]. 海洋环境科学, 23(1): 37–40. LI HONGMEI, MENG HAITAO, 2004. Analysis of environmental factors impacting spatio–temporal variation of benthic community structure in Shenzhen Bay[J]. Marine Environmental Science, 23(1): 37–40 (in Chinese with English abstract).
黎清华, 万世明, 何军, 等, 2014. 近两百年来人类活动对北部湾潮间带环境的影响[J]. 海洋地质与第四纪地质, 34(1): 57–64. LI QINGHUA, WAN SHIMING, HE JUN, et al, 2014. Human impact on the intertidal environment in Beibu Gulf over the last 200 years[J]. Marine Geology & Quaternary Geology, 34(1): 57–64 (in Chinese with English abstract).
李晓静, 周政权, 陈琳琳, 等, 2016. 山东烟台大沽夹河河口及邻近海域大型底栖动物群落特征[J]. 生物多样性, 24(2): 157–165. LI XIAOJING, ZHOU ZHENGQUAN, CHEN LINLIN, et al. Characteristics of macrobenthic communities in the estuary of Dagujia River and its adjacent water areas in Yantai, Shandong[J]. Biodiversity Science, 24(2): 157–165 (in Chinese with English abstract).
刘修泽, 李轶平, 于旭光, 等, 2011. 旅顺南部基岩海岸潮间带大型底栖动物的群落结构研究[J]. 水产科学, 30(12): 777–780. LIU XIUZE, LI YIPING, YU XUGUANG, et al, 2011. The community structure of macrobenthos in intertidal zones in rocky shore in south Lvshun in Dalian[J]. Fisheries Science, 30(12): 777–780 (in Chinese with English abstract).
陆健健, 2003. 河口生态学[M]. 北京: 海洋出版社. LU JIANJIAN, 2003. Estuary ecology[M]. Beijing: China Ocean Press (in Chinese).
许铭本, 赖俊翔, 张荣灿, 等, 2015. 北仑河口北岸潮间带大型底栖动物生态特征及潮间带环境质量评价[J]. 广东海洋大学学报, 35(1): 57–61. XU MINGBEN, LAI JUNXIANG, ZHANG RONGCAN, et al, 2015. Ecological characteristics of macrobenthic animals and environmental quality on the north shore intertidal zone of Beilun estuary[J]. Journal of Guangdong Ocean University, 35(1): 57–61 (in Chinese with English abstract).
叶属峰, 纪焕红, 曹恋, 等, 2004. 河口大型工程对长江河口底栖动物种类组成及生物量的影响研究[J]. 海洋通报, 23(4): 32–37. YE SHUFENG, JI HUANHONG, CAO LIAN, et al, 2004. Studies on the impacts of large-scale estuarine engineering on species composition and biomass of benthos in the Yangtze River estuary[J]. Marine Science Bulletin, 23(4): 32–37 (in Chinese with English abstract).
袁兴中, 陆健健, 2002. 长江口潮滩湿地大型底栖动物群落的生态学特征[J]. 长江流域资源与环境, 11(5): 414–420. YUAN XINGZHONG, LU JIANJIAN, 2002. Ecological characteristics of macrozoobenthic community of tidal flat wetland in the Changjiang estuary[J]. Resources and Environment in the Yangtze Basin, 11(5): 414–420 (in Chinese with English abstract).
张敬怀, 2014. 珠江口及邻近海域大型底栖动物多样性随盐度、水深的变化趋势[J]. 生物多样性, 22(3): 302–310. ZHANG JINGHUAI, 2014. The variation of biodiversity of macrobenthic fauna with salinity and water depth near the Pearl Estuary of the northern South China Sea[J]. Biodiversity Science, 22(3): 302–310 (in Chinese with English abstract).
张莹, 吕振波, 徐宗法, 等, 2012. 环境污染对小清河口大型底栖动物多样性的影响[J]. 生态学杂志, ZHANG YING, LV ZHENBO, XU ZONGFA, et al, 2012, 31(2): 381–387. ZHANG YING, LÜ ZHENBO, XU ZONGFA, et al, 2012. Impacts of environmental pollution on macrobenthos diversity in Xiaoqing estuary of Shandong Province, East China[J]. Chinese Journal of Ecology, 31(2): 381–387 (in Chinese with English abstract).
赵永强, 曾江宁, 高爱根, 等, 2009. 椒江口滩涂大型底栖动物群落格局与多样性[J]. 生物多样性, 17(3): 303–309. ZHAO YONGQIANG, ZENG JIANGNING, GAO AIGEN, et al, 2009. Community pattern and diversity of macrozoobenthos in an intertidal flat, Jiaojiang Estuary[J]. Biodiversity Science, 17(3): 303–309 (in Chinese with English abstract).
周进, 纪炜炜, 2012. 三都澳大型底栖动物次级生产力[J]. 海洋渔业, 34(1): 32–38. ZHOU JIN, JI WEIWEI, 2012. Secondary productivity of macrobenthos in Sandu Bay[J]. Marine Fisheries, 34(1): 32–38 (in Chinese with English abstract).
ARBI I, ZHANG JINGPING, LIU SONGLIN, et al, 2017. Benthic habitat health assessment using macrofauna communities of a sub-tropical semi-enclosed bay under excess nutrients[J]. Marine Pollution Bulletin, 119(2): 39–49.
BELAN T A, 2003. Marine environmental quality assessment using polychaete taxocene characteristics in Vancouver Harbour[J]. Marine Environmental Research, 57(1–2): 89–101.
CARCEDO M C, FIORI S M, PICCOLO M C, et al, 2015. Variations in macrobenthic community structure in relation to changing environmental conditions in sandy beaches of Argentina[J]. Estuarine, Coastal and Shelf Science, 2015, 166: 56-64.
CHAPMAN M G, TOLHURST T J, 2004. The relationship between invertebrate assemblages and bio-dependant properties of sediment in urbanized temperate mangrove forests[J]. Journal of Experimental Marine Biology and Ecology, 304(1): 51–73.
EDGAR G J, BARRETT N S, 2002. Benthic macrofauna in Tasmanian estuaries: Scales of distribution and relationships with environmental variables[J]. Journal of Experimental Marine Biology and Ecology, 270(1): 1–24.
FUJII T, 2007. Spatial patterns of benthic macrofauna in relation to environmental variables in an intertidal habitat in the Humber estuary, UK: Developing a tool for estuarine shoreline management[J]. Estuarine, Coastal and Shelf Science, 75(1–2): 101–119.
GESTEIRA J L G, DAUVIN J-C, 2000. Amphipods are good bioindicators of the impact of oil spills on soft-bottom macrobenthic communities[J]. Marine Pollution Bulletin, 40(11): 1017–1027.
OCCHIPINTI-AMBROGI A, SAVINI D, FORNI G, 2005. Macrobenthos community structural changes off Cesenatico coast (Emilia Romagna, Northern Adriatic), a six-year monitoring programme[J]. Science of the Total Environment, 353(1–3): 317–328.
OTANI S, KOZUKI Y, KURATA K, et al, 2008. Relationship between macrobenthos and physical habitat characters in tidal flat in eastern Seto Inland Sea, Japan[J]. Marine Pollution Bulletin, 57(1–5): 142–148.
PICANÇO T C, ALMEIDA C M R, ANTUNES C, et al, 2014. Influence of the abiotic characteristics of sediments on the macrobenthic community structure of the Minho estuary saltmarsh (Portugal)[J]. Limnetica, 33(1): 73–88.
REHITHA T V, ULLAS N, VINEETHA G, et al, 2017. Impact of maintenance dredging on macrobenthic community structure of a tropical estuary[J]. Ocean & Coastal Management, 144: 71–82.
SARKER J, TANMAY M H, RAHMAN F, et al, 2016. Assessment of coastal water pollution in Greater Noakhali-Bangladesh[J]. Journal of Coastal Zone Management, 19: 427.
Study on the macrozoobenthic community structure in intertidal zone of Fangchenghe Estuary of Guangxi, China
LAI Tinghe1, 2, HE Binyuan1, 2, HUANG Zhongjian2, TANG Qiao3, QIN Luyan2, ZHU Ting2, MO Zhenni2, LIU Li2, ZHONG Yunxu4
1. QinzhouUniversity, Qinzhou 535011, China; 2. Guangxi Academy of Oceanography, Nanning 530022, China; 3. Guangxi Beilun Estuary National Reserve, Fangchenggang 538100, China; 4. Guangxi Mangrove Research Center, Beihai 536000, China
Based on the survey in May, August and November 2016 and February 2017, the effects of natural and anthropogenic factors on macrozoobenthic community structure in the intertidal zone of Fangchenghe Estuary of Guangxi, China were studied. A total of 252 species belonging to 10 phyla of intertidal macrozoobenthos were collected, among which Mollusca, Arthropoda and Annelida were richer in species abundance.,,,,,,, andwere ranked as the top eight dominant species in this study area through the whole year. The annual average density and biomass of the study area were 203 ind×m-2and 276.58 g×m-2, respectively, in which Mollusca occupied 69.58% and 83.73% to the total, respectively. ANOVA showed that salinity and sediment type factors had significant influences (<0.05 and less) on macrozoobenthos density, biomass, species diversity′, Richness, Evenness, and quadrat-based species abundance, while season and tidal zone factors had very small influences (>0.05) on them. Salinity had larger influences on density and biomass than sediment type did, while sediment type had larger influences on species diversity′, Richness, Evenness, and quadrat-based species abundance than salinity did. Both clustering analysis and multidimensional scaling analysis obtained the same results that the 21 sampling stations could be divided into three groups, basically owing to the effects from salinity and sediment type. The status of diversity index′ showed that in general the intertidal macrozoobenthic communities were under middle-level fluctuation. Comprehensively, three factors, i.e., salinity, sediment type and anthropogenic fluctuation, played combined roles on the spatial distribution pattern of macrozoobenthic communities in the intertidal zone of Fangchenghe Estuary.
Fangchenghe Estuary; intertidal macrozoobenthos; community structure; environmental fluctuation
2018-05-31;
2018-08-29. Editor: YIN Bo
Science and Technology Project of State Oceanic Administration (YLFCJ20164006-F); Science and Technology Project of Guangxi Oceanic Administration (GXHYJ100); National Key Research and Development Program of China (2017YFC0506100)
HE Binyuan. Email: hebinyuan2008@126.com
P735.542
A
1009-5470(2019)02-0067-11
2018-05-31;
2018-08-29。殷波编辑
国家海洋局专项(YLFCJ20164006-F); 广西海洋局研究专项(GXHYJ100); 国家重点研发计划重点专项(2017YFC0506100)
赖廷和(1970—), 女, 广西壮族自治区合浦县人, 副研究员, 主要从事海洋生态和海洋环境研究。E-mail: laitinghe@163.com
何斌源。Email: hebinyuan2008@126.com
10.11978/2018058
