褐菖鲉仔、稚鱼消化系统发育的组织学观察
2019-04-11杨佳喆齐闯徐善良
杨佳喆, 齐闯, 徐善良
褐菖鲉仔、稚鱼消化系统发育的组织学观察
杨佳喆1, 齐闯1, 徐善良2
1. 宁波大学海洋学院, 浙江 宁波 315211; 2. 应用海洋生物技术教育部重点实验室, 宁波大学, 浙江 宁波 315211
为探究卵胎生鱼类褐菖鲉()的仔、稚鱼消化系统组织学特点, 采用常规组织切片技术, 并用HE染色和组织化学染色, 比较分析了产出后0~50d的褐菖鲉仔、稚鱼消化系统发育的组织学变化。结果显示: 褐菖鲉初产仔鱼已分化出口咽腔, 并具有初始的食道、胃、肠、肝脏和胰脏; 2日仔鱼肛门与外界相通, 食道扩大, 开始摄食, 进入混合性营养期; 3~4日仔鱼幽门盲囊出现, 食道内壁出现黏膜皱褶5~6个, 胃黏膜皱褶5个, 肝细胞团区域扩大; 5~6日仔鱼卵黄囊和油球耗尽, 进入外源性营养期, 食道黏膜上皮出现杯状细胞, 肠道弯曲, 可区分前中肠和后肠两部分; 10~14日仔鱼食道环肌层明显, 黏膜皱褶增加到7~12个, 胃壁四层结构基本形成, 肠道黏膜皱褶加深, 纹状缘清晰可见, 肝细胞分裂迅速, 数量显著增加, 体积增大, 出现肝血窦和胰岛; 28~30日稚鱼出现胃腺和胃小凹, 已具功能性胃, 标志着稚鱼期的开始, 此时胰岛细胞数量和酶原颗粒增多, 消化能力显著提高; 47~50日稚鱼已基本具有成鱼胃的特征, 肠道纹状缘发达, 肝细胞呈多边形, 细胞内含大量脂肪颗粒, 消化系统从结构和功能上已趋于完善。由此可见, 褐菖鲉仔鱼和稚鱼消化系统的发育具有卵胎生鱼类发育较早的特点, 与其消化功能的完善密切相关。
褐菖鲉; 仔稚鱼; 消化系统; 组织学
褐菖鲉(), 俗称“石头鲈”、“虎头鱼”等, 隶属于鲉形目(Scorpaeniformes)、鲉亚目(Scorpaenoidei)、鲉科(Scorpaenidae)和菖鲉属(), 是一种底层肉食性卵胎生鱼类。广泛分布于中国、朝鲜半岛、日本、菲律宾等国近海岩礁海域, 也是开展人工养殖与增殖放流的优良海水鱼种之一(石戈等, 2007)。
鱼类消化系统结构决定了鱼类的食性(徐革锋等, 2009), 研究消化系统的结构和功能是认识和探讨鱼类消化生理的基础和途径之一(方华华等, 2011), 是作为监测鱼类幼苗生长发育过程中消化能力和营养需求变化规律的一种常用手段, 可进一步优化幼苗的培育条件(Micale et al, 2010)。国内外对多种卵生硬骨鱼类仔、稚、幼鱼消化系统发育的研究已有较多报道, 如北美牙鲆()(Gisbert et al, 2004)、银鲳()(高露姣等, 2007)、大黄鱼()(徐晓津等, 2010)、卵形鲳鲹()(区又君等, 2011)、黄鳍鲷()(王永翠等, 2012)、条石鲷()(区又君等, 2015)、大西洋白姑鱼()(Solovyev et al, 2016)等, 而对卵胎生硬骨鱼类的研究较少, 仅见林强等(2007)对大海马()消化系统早期发育的组织学特征的研究。
近几年有关褐菖鲉的国内外研究, 主要涉及褐菖鲉成鱼的消化道组织学和组织化学特性(石戈等, 2007), 褐菖鲉脑部结构特性(Murakami et al, 1983, 2010)和环境污染物对其的影响(Zheng et al, 2016; 孙文静等, 2018)等方面, 对褐菖鲉仔、稚鱼的消化系统组织学研究目前仍为空白, 本文探讨了褐菖鲉消化机能的早期发育规律, 为人工育苗中饵料的供给提供参考。
1 材料和方法
1.1 实验用鱼
从温州市洞头区海域捕获的野生成熟褐菖鲉中挑选实验用褐菖鲉亲鱼, 在浙江省海洋水产养殖研究所洞头基地水泥养殖池中自然分娩产出的仔鱼。将同一天产出的仔鱼及时转移到育苗池培育, 1~10d仔鱼投喂轮虫, 10~15d仔鱼混合投喂轮虫和丰年虫, 15~20d仔鱼后投喂丰年虫和桡足类, 28d进入稚鱼期后, 投喂桡足类为主, 35~50d稚鱼增加投喂鱼、虾穈和配合饲料。
培养水温为15~17℃, 盐度为27‰~32‰, pH为7.8~8.2, 溶氧在6mg×L-1以上, 每天换水1次。
1.2 取样固定及组织切片的制作
1~20d每天取样, 20d后隔2~3d取样, 取样持续至50d稚鱼。于每日早上投喂前取样, 每次取样30尾左右。样品用Bouin’s氏液固定24h后, 用70%乙醇溶液洗脱保存。经常规脱水透明、石蜡包埋等程序处理后, 分别对组织块进行横、纵连续切片, 切片厚度为6μm, 常规HE染色(金国雄等, 2013)。
对23日仔鱼、30日和33日稚鱼样品进行组织化学研究。样品用Carnoy氏液固定, 采用汞-溴酚蓝法显示蛋白质(Pearse, 1983)。
所有切片在Olympus-BX51显微镜下观察, 并选取典型结构进行拍照。
2 结果
2.1 褐菖鲉的消化道发育
2.1.1 口咽腔
初产的褐菖鲉仔鱼, 口咽腔已形成, 具有一层口腔黏膜上皮细胞, 此时无黏膜下层和肌肉层的出现。2日仔鱼的口咽腔扩大, 上颌口腔瓣和下颌口腔瓣都已出现, 上颌比下颌长(图1b)。3日时, 口咽腔出现肌肉层, 此时的口咽腔结构已具有黏膜层、黏膜下层和肌肉层(图1c)。10日仔鱼的口咽腔后部明显增厚并出现皱褶, 同时黏膜上皮出现杯状细胞和黏液细胞(图1e)。随后至稚鱼期, 口咽腔的变化集中在杯状细胞和黏液细胞数量的增加上。
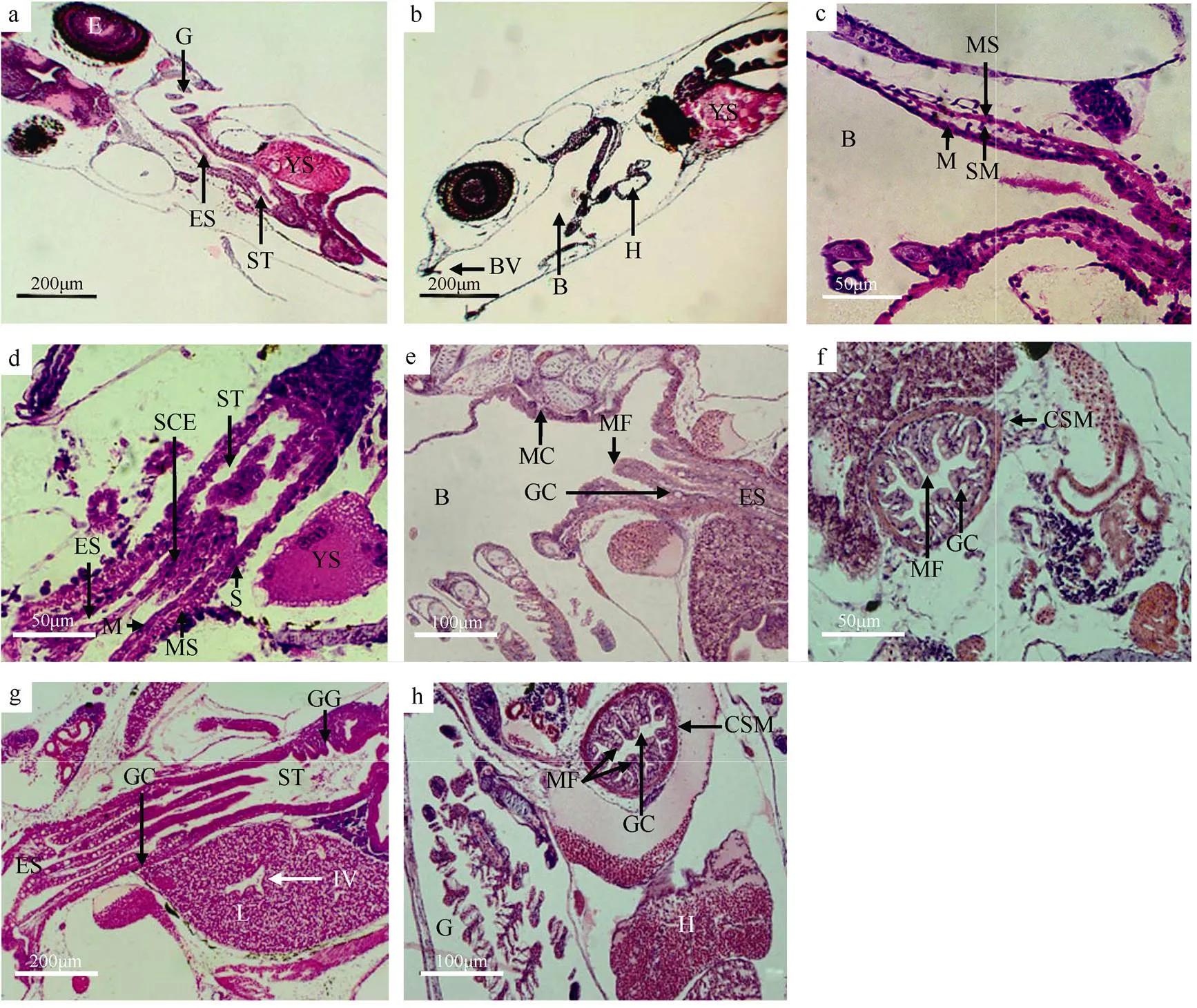
图1 褐菖鲉仔、稚鱼口咽腔和食道发育的组织结构
a. 初产仔鱼消化道整体结构纵切面; b. 2日仔鱼口咽腔整体结构纵切面; c. 3日仔鱼口咽腔结构纵切面; d. 4日仔鱼食道整体结构纵切面; e. 10日仔鱼食道的杯状细胞纵切面; f. 21日仔鱼食道的环层肌和黏膜皱褶横切面; g. 28日稚鱼食道和胃纵切面; h. 39日稚鱼食道横切面。B: 口咽腔; BV: 口咽瓣; CSM: 环肌层; E: 眼; ES: 食道; G: 鳃; GC: 杯状细胞; GG: 胃腺; H: 心脏; IV: 小叶间静脉; L: 肝; M: 黏膜; MC: 黏液细胞; MF: 黏膜褶; MS: 肌肉层; S: 浆膜层; SCE: 单层立方上皮; SM: 黏膜下层; ST: 胃; YS: 卵黄囊
Fig. 1 Histological observation of the development of buccopharyngeal cavity and esophagus of larva and juvenile of.. (a) longitudinal section of digestive tract of larva; (b) longitudinal section of buccopharyngeal cavity of 2 dpp larva; (c) longitudinal section of buccopharyngeal cavity of 3 dpp larva; (d) longitudinal section of esophagus of 4 dpp larva; (e) longitudinal section of goblet cells in esophagus of 10 dpp larva; (f) transverse section of circular layers of striated muscle and mucosal folds in esophagus of 21 dpp larva; (g) longitudinal section of esophagus and stomach of 28 dpp juvenile; (h) transverse section of esophagus of 39 dpp juvenile. B: Buccopharyngeal cavity; BV: buccopharyngeal valve; CSM: circular layers of striated muscle; E: eye; ES: esophagus; G: gill; GC: goblet cell; GG: gastric gland; H: heart; IV: interlobular vains; L: liver; M: mucosa; MC: mucus cell; MF: mucosal fold; MS: muscle layer; S: serosa; SCE: simple columnar epithelium; SM: submucosa; ST: stomach; YS: yolk sac
2.1.2 食道
初产仔鱼已经分化出一个短而狭窄的食道腔, 与初始的胃腔相连, 呈直管状, 具单层细胞的黏膜层, 细胞核为球形, 未出现肌层和杯状细胞(图1a)。产出2日后, 食道逐步扩大, 食道壁也渐增厚。4日仔鱼食道的内层具有复层扁平上皮, 食道到胃过渡带为单层立方上皮, 此时食道内壁出现细小的向内腔纵向凸起的纵行黏膜皱褶5~6个, 较薄的肌肉层也随之发生, 食道组织分为了浆膜层、肌肉层和黏膜层3层结构(图1d)。6日仔鱼的食道黏膜上皮出现杯状细胞, 至10日仔鱼的食道黏膜上皮中有一些黏液细胞出现, 纵行黏膜皱褶增加到7~9个, 肌肉层增厚(图1e)。13日仔鱼食道环肌层明显, 纵行黏膜皱褶增至11~12个。21日仔鱼食道的环肌层发达, 厚度达到7.5μm (图1f)。28日仔鱼食道前段富含大量杯状细胞, 纵肌层仍较薄, 黏液细胞数量大量增加(图1g)。39日稚鱼食道内壁纵行黏膜皱褶达到15个, 肌肉层发达, 环肌层的厚度增至为12.7μm (图1h)。
2.1.3 胃
初产仔鱼已发现原始的胃腔结构, 胃壁与食道的上皮结构相似, 为简单的单层细胞(图2a)。3日仔鱼胃黏膜皱褶5个, 皱褶最高为23μm, 已有分层结构, 但分化不明显(图2b)。4日仔鱼胃的幽门部出现几个管状突出物, 即为分化的幽门盲囊, 以此区分贲门胃、胃体与幽门胃(图2c)。10日仔鱼的胃壁基本成型, 由浆膜层、肌肉层、黏膜下层和黏膜层组成, 胃腔内黏膜纵褶数量增多, 纵褶高35μm, 黏膜层由单皮柱状细胞构成, 细胞紧密排列, 细胞核位于细胞中下部, 黏膜层与黏膜下层较厚, 肌层尚不明显(图2e)。23日仔鱼胃壁肌层增厚, 黏膜褶皱增加(图2h)。28日仔鱼胃黏膜下层中出现少量实心细胞团的胃腺(图1g)。30日仔鱼胃腔内的皱褶加粗, 组织化学显示, 胃壁被汞-溴酚蓝染成深蓝色(图3), 黏膜层和肌肉层染色较深, 黏膜下层和浆膜层则染色稍浅。39日稚鱼胃的组织结构发育更完善, 胃壁出现胃小凹, 胃腺数量增多, 胃黏膜皱褶厚度达90μm (图2j)。47日稚鱼胃中管状胃腺十分丰富, 皱褶大量增多, 黏膜下层和肌肉层增厚, 胃腔进一步扩大, 基本具有成鱼胃的特征(图2k)。
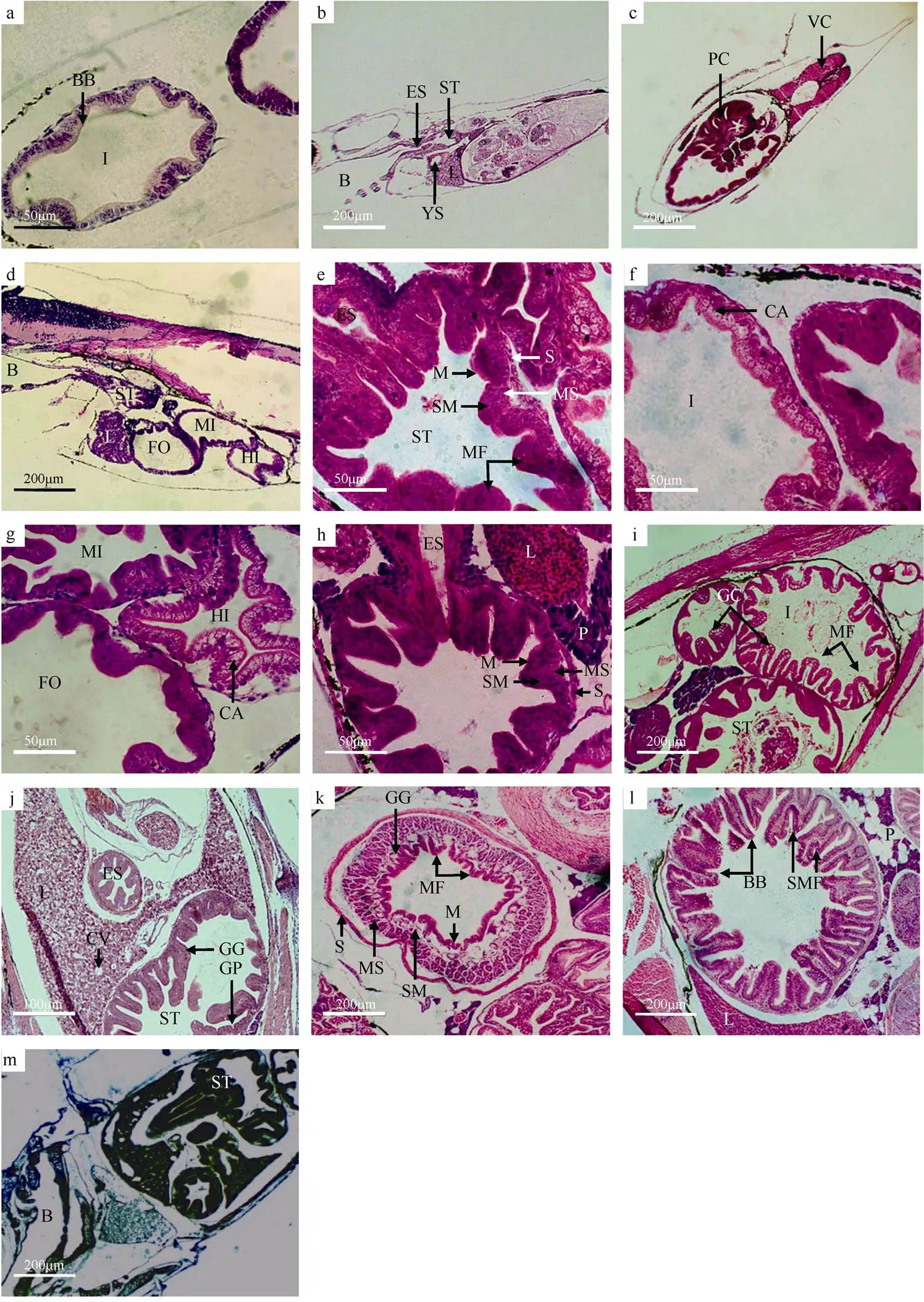
图2 褐菖鲉仔、稚鱼胃和肠发育的组织结构
a. 初产仔鱼肠纹状缘结构横切面; b. 3日仔鱼肝和胃纵切面; c. 4日仔鱼幽门盲囊横切面; d. 8日仔鱼肠道整体结构纵切面; e. 10日仔鱼胃整体结构横切面; f. 10日仔鱼肠上皮细胞空泡结构横切面; g. 13日仔鱼前肠、中肠、后肠横切面; h. 23日仔鱼胃整体结构横切面; i. 28日稚鱼肠整体结构横切面; j. 39日稚鱼胃整体结构横切面; k. 47日稚鱼胃整体结构横切面; l. 47日稚鱼肠道整体结构横切面; m. 30日稚鱼整体纵切面, 汞-溴酚蓝染色。B: 口咽腔; BB: 纹状缘; CA: 空泡; CV: 中央静脉; ES: 食道; FO: 前肠; GC: 杯状细胞; GP: 胃小凹; HI: 后肠; I: 肠; L: 肝; M: 黏膜; MF: 黏膜褶; MI: 中肠; MS: 肌肉层; P: 胰; PC: 幽门盲囊; S: 浆膜层; S: 浆膜层; SM: 黏膜下层; SMF: 次级黏膜褶; VC: 脊椎; YS: 卵黄囊
Fig. 2 Histological observation of the development of stomach and intestine of larva and juvenile of.. (a) transverse section of brush border in intestine of larva; (b) longitudinal section of liver and stomach of 3 dpp larva; (c) transverse section of pyloric caecum of 4 dpp larva; (d) longitudinal section of intestine of 8 dpp larva; (e) transverse section of stomach of 10 dpp larva; (f) transverse section of cavitation in intestinal epithelium of 10 dpp larva; (g) transverse section of foregut, hindgut and midgutof 13 dpp larva; (h) transverse section of stomach of 23 dpp larva; (i) transverse section of intestine of 28 dpp juvenile; (j) transverse section of stomach of 39 dpp juvenile; (k) transverse section of stomach of 47 dpp juvenile; (l) transverse section of intestine of 47 dpp juvenile; (m) longitudinal section of stomach of 30 dpp juvenile by Mercury Bromophenol blue method. B: Buccopharyngeal cavity; BB: brush border; CA: cavitation; CV: central vein; ES: esophagus; FO: foregut; GC: goblet cell; GP: gastric pit; HI: hindgut; I: intestine; L: liver; M: mucosa; MF: mucosal fold; MI: midgut; MS: muscle layer; P: pancreas; PC: pyloric caecum; S: serosa; SM: submucosa; SMF: Secondary mucosal fold; VC: vertebral column; YS: yolk sac
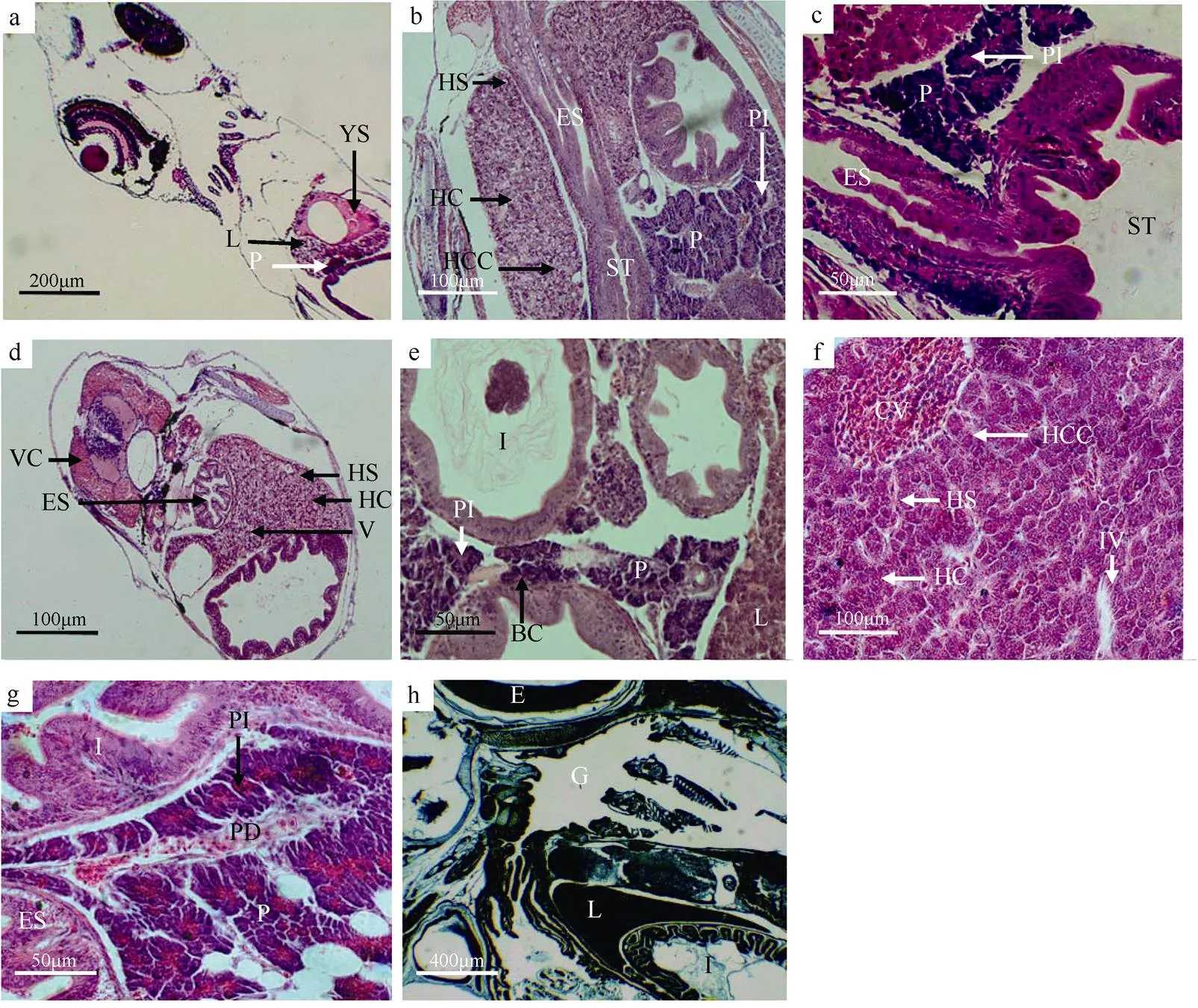
图3 褐菖鲉仔、稚鱼肝脏和胰脏发育的组织结构
a. 初产仔鱼消化腺整体结构纵切面; b. 10日仔鱼肝脏和胰脏整体结构纵切面; c. 13日仔鱼胰脏整体结构纵切面; d. 14日仔鱼肝脏横切面; e. 20日仔鱼胰脏横切面; f. 47日稚鱼肝脏整体结构纵切面; g. 50日稚鱼胰脏整体结构横切面; h. 23日仔鱼整体纵切面, 汞-溴酚蓝染色。B: 口咽腔; BC: 血细胞; CV: 中央静脉; E: 眼; ES: 食道; G: 鳃; HC: 肝细胞; HCC: 肝细胞索; HS: 肝血窦; I: 肠; IV: 小叶间静脉; L: 肝; P: 胰脏; PD: 胰管; PI: 胰岛; ST: 胃; V: 空泡结构(肝); VC: 脊椎; YS: 卵黄囊
Fig. 3 Histological observation of the development of liver and pancreas of larva and juvenile of.. (a) longitudinal section of digestive gland of larva; (b) longitudinal section of liver and pancreas of 3 dpp larva; (c) longitudinal section of pancreas of 13 dpp larva; (d) transverse section of liver of 14 dpp larva; (e) transverse section of pancreas of 20 dpp larva; (f) longitudinal section of liver of 47 dpp juvenile; (g) transverse section of pancreas of 50 dpp juvenile; (h) longitudinal section of liver of 23 dpp larva by Mercury Bromophenol blue method. B: Buccopharyngeal cavity; BC: blood cell; CV: central vein; E: eye; ES: esophagus; G: gill; HC: hepatic cell; HCC: hepatic cell cords; HS: hepatic sinusoid; I: intestine; IV: interlobular vains; L: liver; P: pancreas; PD: pancreatic duct; PI: pancreatic island; ST: stomach; V: vacuolar structure (liver); VC: vertebral column; YS: yolk sac
2.1.4 肠
初产仔鱼尚未摄食, 但已出现肠的结构, 由单层柱状细胞组成, 细胞核位于中部, 靠近肠腔的内层可见HE染色呈淡红色的纹状缘, 出现低矮褶皱(图2a)。2日仔鱼的肛门与外界连通, 肠道中已发现食物。5日仔鱼肠道弯曲, 可区分前中肠和后肠两部分。至8日, 肠道结构完整, 也由浆膜层、肌肉层、黏膜下层和黏膜层组成, 肠道出现2个肠曲, 使前肠、中肠和后肠界限清晰, 肠黏膜细胞上皮及黏膜皱褶高度明显增加(图2d)。10日仔鱼肠上皮细胞出现空泡结构(图2f)。13日仔鱼后肠染色较浅, 细胞核位于细胞底部, 纹状缘清晰可见, 黏膜皱褶继续加深, 后肠皱褶比前中肠更甚(图2g)。21日仔鱼肠上皮细胞核上空泡数量增加。28日仔鱼肠道的单层柱状细胞排列更加规则、紧密, 细胞核位于细胞中下部; 肠道黏膜上皮的固有膜和黏膜下层界限不明显, 黏膜上皮分布许多杯状细胞, 排列无规律; 黏膜皱褶丰富, 皱褶的排列方向不一, 肌层增厚(图2i)。47日稚鱼, 肠道纹状缘发达, 黏膜皱褶更丰富, 同时出现次级皱褶(图2l)。
2.2 消化腺发育
2.2.1 肝脏
初产仔鱼在卵黄囊和消化道之间出现略呈三角形、染色较浅且结构松散的肝细胞团, 即为肝脏原基, 其细胞界限不是很清晰, 在肝脏与卵黄囊接触的反侧, 卵黄液化区域内可见一些未成熟的血细胞(图3a)。3日仔鱼的肝脏细胞团依三角形区域扩大, 细胞核大, 位于肝细胞中央(图2b)。10日仔鱼肝细胞分裂迅速, 数量显著增加, 肝脏体积增大。肝细胞染色加深, 呈索状排列, 已出现大量空泡结构, 细胞核被挤到一侧, 即肝细胞开始贮存脂质。同时出现肝血窦, 数量多但体积小, 肝血窦内有少许未发育完全的血细胞(图3b)。14日仔鱼肝细胞排列十分致密, 空泡结构大量增多, 肝血窦数量增多(图3d)。23日仔鱼, 组织化学结果显示, 肝组织被汞-溴酚蓝染成深蓝色, 细胞核和细胞质染色深度一致(图3h)。28日仔鱼肝血窦内有大量血细胞, 肝脏血管丰富。47日稚鱼肝细胞多呈多边形结构, 核染色深呈球形, 位于细胞一侧, 细胞内含大量脂肪颗粒, 不易染色, 呈透亮状。由肝细胞排列而成的肝细胞索以中央静脉为中心, 向周围呈辐射状分布, 此时肝脏在组织学上与成体一致(图3f)。
2.2.2 胰脏
初产仔鱼在肝的下方出现紫蓝色的胰腺细胞团, 且有少部分已埋于肝腹侧(图3a)。7日仔鱼胰腺细胞体积稍有增大, 已经聚集为胰脏。10日仔鱼可见散布在外分泌部中的胰岛, 其细胞颜色较浅, 呈圆团状分布, 排列疏松, 血细胞较少(图3b)。13日仔鱼胰腺细胞分化速度快, 细胞数量增加, 为不规则结构, 核为圆形。细胞之间界限不清晰, 含有大量酶源颗粒。胰腺细胞主要分布于肝与胃之间, 颜色加深, 细胞核位于细胞中央(图3c)。20日仔鱼, 胰岛增多, 胰脏静脉血管明显, 分布少量血细胞(图3e)。28日仔鱼胰岛细胞数量增加, 体积进一步扩大, 酶原颗粒增多, 间隙增大。50日稚鱼胰腺发育更加完全, 在组织学上和成体结构基本一致(图3g)。
3 讨论
3.1 发育水温与仔、稚鱼消化系统发育的关系
口和肛门的形成及其与外界相通是仔鱼即将开口摄食的重要标志之一(Anderson et al, 2012)。有研究表明, 在适温范围内, 水温越高仔鱼口和肛门与外界相通以及卵黄囊耗尽的时间越早(刘鉴毅等, 2015; Faccioli et al, 2016)。在中国东海海域, 褐菖鲉的生殖季节为11月至翌年4月间, 此时的自然水温为10~18℃(林丹军等, 2002; 邱成功等, 2013), 本研究褐菖鲉的培育水温为15~17℃, 处于其自然繁殖水温范围内。
有胃硬骨鱼类的消化系统早期发育主要可分为四个时期: 从仔鱼孵出或产出到其开口摄食的阶段为内源性营养期; 仔鱼开口摄食外源性食物到其卵黄完全吸收期间, 则被称作内源和外源混合性营养期; 仔鱼完全依赖外源性食物的外源性营养期 Ⅰ (不具功能性胃)以及胃腺出现后稚鱼期开始的外源性营养期 Ⅱ (具有功能性胃)(Faccioli et al, 2016)。本研究发现, 褐菖鲉的消化系统发育亦具有上述阶段性特征。几种有胃及无胃硬骨鱼类的消化系统早期发育时期比较见表1。
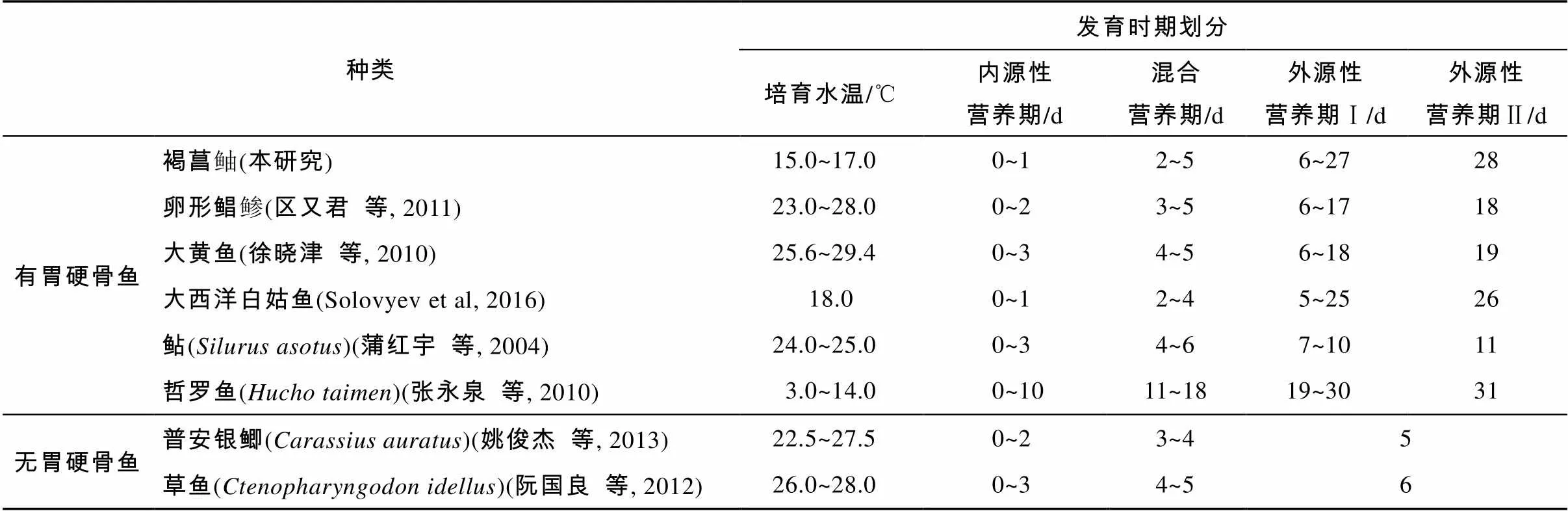
表1 几种有胃及无胃硬骨鱼类的消化系统早期发育时期划分
由表1可知, 普安银鲫和草鱼等鲤科鱼类属无胃鱼类, 与多数有胃鱼类不同, 其消化系统发育的差异直接导致了其仔稚鱼发育阶段划分的差异(谢木娇等, 2017)。此外, 种间不同的发育水温也直接影响着鱼类消化系统的发育速度。培育水温较低的大西洋白姑鱼和哲罗鱼等鱼类进入具有功能性胃的外源性营养期Ⅱ 的时间需要26~31d, 远比较高温度的大黄鱼等鱼类时间长, 即水温低鱼类的消化系统发育慢, 主要表现为胃腺出现的时间较晚。本研究中的褐菖鲉虽为卵胎生繁殖类型, 但也具有该发育特性。
3.2 褐菖鲉仔、稚鱼消化系统发育的特点
褐菖鲉属海洋中少数的卵胎生鱼类之一, 孵化后需在卵巢液中生活几天后才产出体外, 故初产仔鱼的口咽腔已形成, 这与卵生硬骨鱼类都不同。林丹军等(2002)认为褐菖鲉仔鱼在母体内孵化后第5日口部初开, 7日产出。邱成功等(2013)也发现1日仔鱼上下颌已经形成, 且偶尔可见下颌活动。而大海马仔鱼产出后即可独立摄食, 在育儿囊内完成了内源性营养期, 直接进入混合营养期(林强等, 2007)。此外, 褐菖鲉初产仔鱼还具有原始的食道, 胃腔、肠结构, 以及独立的肝、胰脏, 1日龄仔鱼的消化系统明显比大黄鱼(徐晓津等, 2010)等大多数海水鱼类发育快。褐菖鲉和大海马的这种现象, 与其在亲代体内发育了一段时间再产出有关。褐菖鲉初产仔鱼虽然开口, 但尚未摄食, 其所需营养物质与所有硬骨鱼类一样, 仍由其卵黄囊提供。
褐菖鲉2日仔鱼已能摄食外源性食物(邱成功等, 2013)。辅助消化器官幽门盲囊分化也较早, 这与大黄鱼(徐晓津等, 2010)等大多数有胃硬骨鱼类在胃腺出现前后才分化不同。幽门盲囊与食道和胃出现的黏膜皱褶, 可以扩大消化道的贮存容积和消化面积, 利于食物的充分消化和吸收(勾效伟等, 2008; 区又君等, 2015)。相关研究都表明, 海水鱼仔鱼开口摄食后的第一周是至关重要的阶段, 特别是在外源性摄食开始和卵黄囊吸收完成时(Solovyev et al, 2016), 如不及时供给适合开口饵料将会影响到仔鱼生长甚至造成死亡(Sarasquete et al, 1995)。对褐菖鲉而言, 2~5日是内源性转向外源性营养的关键阶段, 此时要保障适口的轮虫供给。
褐菖鲉的卵黄囊持续时间与大多数有胃硬骨鱼类相似(表1), 5日褐菖鲉仔鱼的卵黄囊和油球完全吸收, 进入外源性营养期Ⅰ。6日后仔鱼的食道复层上皮出现杯状细胞, 表明能够分泌中性和酸性黏液, 在摄食食物时, 可以更好地润滑和保护上皮组织, 起到保护上皮组织减少摩擦和避免伤害的作用(Díaz et al, 2008; Faccioli et al, 2016)。而中性黏液起到乳化食物变成食糜的作用(Murray et al, 1996), 也证明了褐菖鲉的消化作用可能在食道就开始了, 尤其是对通过咽齿咀嚼后形成的一些小颗粒食物的消化(石戈等, 2007)。8~10日仔鱼的胃基本成型, 肠道结构基本完整, 可明显区分前肠、中肠和后肠, 其肠黏膜皱褶初具成体特性: 后肠皱褶最密, 呈网状褶; 中肠皱褶最高, 呈纵行褶, 而前肠皱褶的密度和高度都低于前肠和中肠(石戈等, 2007)。10日仔鱼肠上皮细胞出现空泡, 揭示了仔鱼发育期间, 消化道存在着胞吞和细胞内消化作用(区又君等, 2015)。同时, 肝脏出现大量空泡结构, 这些空泡为储存在肝脏中的糖原(Boulhic et al, 1992)。组织化学实验结果显示, 23日肝脏中存储的蛋白质含量已很高。许多研究表明, 硬骨鱼类肝脏和胰脏发育时间存在种属间的差异。褐菖鲉肝脏和胰脏皆为独立器官, 与具有弥散性胰腺的卵形鲳鲹(区又君等, 2011)等鱼类不同, 而与大黄鱼(徐晓津等, 2010)等一致。褐菖鲉初产仔鱼已同时具有肝脏和胰脏, 林丹军等(2002)研究发现在母体内孵化后第3日仔鱼的肠管旁已出现肝脏组织, 早于胰脏出现, 次序上与大部分鱼类一致。仔鱼外源性摄食的发生, 主要依赖于胰腺分泌的消化酶, 褐菖鲉仔鱼拥有一个在形态和功能上发育比较好的胰腺, 为其开口摄食提供了良好的保障。在外源性营养期 Ⅰ (胃前消化), 褐菖鲉仔鱼主要依靠黏液细胞和杯状细胞的分泌物, 通过肠道和胰腺中的酶类进行消化, 经由肠上皮细胞吸收到达淋巴细胞和血液, 最后存储在肝脏中。
28日褐菖鲉的胃出现胃腺, 其胃腺出现时间较晚。组织化学实验证实在胃腺出现2d (产出30d)后胃中的蛋白含量就明显增多, 表明了胃腺大量分泌胃蛋白酶消化外源性食物蛋白。功能性胃的出现, 标志着褐菖鲉稚鱼期的开始和更完善的消化机制的形成(Tanaka, 1971; Sarasquete et al, 1995; Solovyev et al, 2016)。这比邱成功等(2013)在形态学上以35d作为仔鱼与稚鱼期的划分时间提前了一周左右, 该现象也在条石鲷(区又君等, 2015)和青龙斑(李加儿等, 2016)等硬骨鱼类中存在。同时, 在褐菖鲉的胃黏膜上皮中没有发现消化道常见的杯状细胞, 揭示了其胃消化功能主要依赖于强大的胃腺(石戈等, 2007)。与此同时, 食道和肠道的主要变化是皱褶的加深和肌肉层的增厚, 而幽门盲囊结构也更趋于完整。47~50日褐菖鲉稚鱼的消化系统在质的方面已于成鱼基本一致, 随着鳞被形成, 进入幼鱼期(邱成功等, 2013)。
综上所述, 褐菖鲉消化系统的发育具有卵胎生鱼类发育较早的特性, 也符合有胃硬骨鱼类消化系统发育的规律, 体现了褐菖鲉消化系统的发育与其功能相一致的关系。
方华华, 王波, 2011. 星斑川鲽仔稚鱼消化系统发育的组织学研究[J]. 中国农学通报, 27(14): 50–54. FANG HUAHUA, WANG BO, 2011. Histological studies on the development of digestive system in larval and juvenile Starry flounder[J]. Chinese Agricultural Science Bulletin, 27(14): 50–54 (in Chinese with English abstract).
高露姣, 施兆鸿, 严莹, 2007. 银鲳仔鱼消化系统的组织学研究[J]. 中国水产科学, 14(4): 540–546. GAO LUJIAO, SHI ZHAOHONG, YAN YING, 2007. Histological studies on development of digestive system in larval[J]. Journal of Fishery Sciences of China, 14(4): 540–546 (in Chinese with English abstract).
勾效伟, 区又君, 廖锐, 2008. 平鲷消化系统形态学、组织学及组织化学研究[J]. 南方水产, 4(5): 28–36. GOU XIAOWEI, OU YOUJUN, LIAO RUI, 2008. Histological and histochemical study of digestive system of[J]. South China Fisheries Science, 4(5): 28–36 (in Chinese with English abstract).
金国雄, 徐伟, 耿龙武, 等, 2013. 大鳞鲃消化系统早期发育的组织学观察[J]. 水产科学, 32(6): 311–315. JIN GUOXIONG, XU WEI, GENG LONGWU, et al, 2013. Histological observation of early ontogenetic development of digestive system in Bulatmail barbel[J]. Fisheries Science, 32(6): 311–315 (in Chinese with English abstract).
李加儿, 吴水清, 区又君, 等, 2016. 斜带石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代(青龙斑)消化系统的早期发育[J]. 动物学杂志, 51(1): 73–83. LI JIAER, WU SHUIQING, OU YOUJUN, et al, 2016. Post-embryonic development of the digestive system in Qinglong grouper (♀×♂)[J]. Chinese Journal of Zoology, 51(1): 73–83 (in Chinese with English abstract).
林丹军, 尤永隆, 2002. 卵胎生硬骨鱼褐菖鲉胚胎及仔鱼的发育[J]. 台湾海峡, 21(1): 45–52. LIN DANJUN, YOU YONGLONG, 2002. Embryonic and larval development of ovoviviparous teleost,[J]. Journal of Oceanography in Taiwan Strait, 21(1): 45–52 (in Chinese with English abstract).
林强, 吕军仪, 张彬, 等, 2007. 大海马消化系统胚后发育的形态学及组织学研究[J]. 热带海洋学报, 26(6): 46–51. LIN QIANG, LÜ JUNYI, ZHANG BIN, et al, 2007. Histological studies on post-embryonic development of digestive system of Seahorse[J]. Journal of Tropical Oceanography, 26(6): 46–51 (in Chinese with English abstract).
刘鉴毅, 宋志明, 王妤, 等, 2015. 温度对点篮子鱼幼鱼生长、摄食和消化酶活性的影响[J]. 海洋渔业, 37(5): 442–448. LIU JIANYI, SONG ZHIMING, WANG YU, et al, 2015. Effects of water temperature on growth, feeding and activities of digestive enzymes of juvenile[J]. Marine Fisheries, 37(5): 442–448 (in Chinese with English abstract).
区又君, 何永亮, 李加儿, 2011. 卵形鲳鲹消化系统的胚后发育[J]. 台湾海峡, 30(4): 533–539. OU YOUJUN, HE YONGLIANG, LI JIAER, 2011. Postembryonic development of digestive system of[J]. Journal of Oceanography in Taiwan Strait, 30(4): 533–539 (in Chinese with English abstract).
区又君, 李加儿, 艾丽, 2015. 条石鲷早期发育阶段消化系统组织学研究[J]. 华南农业大学学报, 36(1): 23–27. OU YOUJUN, LI JIAER, AI LI, 2015. A study on the histology of digestive system in early life stages of[J]. Journal of South China Agricultural University, 36(1): 23–27 (in Chinese with English abstract).
蒲红宇, 翟宝香, 刘焕亮, 2004. 鲇仔、稚鱼消化系统胚后发育的组织学观察研究[J]. 中国水产科学, 11(1): 1–8. PU HONGYU, ZHAI BAOXIANG, LIU HUANLIANG, 2004. Histological studies on post-embryonic development of digestive system in larval catfish[J]. Journal of Fishery Sciences of China, 11(1): 1–8 (in Chinese with English abstract).
邱成功, 徐善良, 齐闯, 等, 2013. 褐菖鲉()早期生长发育与人工繁育技术研究[J]. 宁波大学学报(理工版), 26(4): 17–23. QIU CHENGGONG, XU SHANLIANG, QI CHUANG, et al, 2013. Studies on early growth and development inwith artificial breeding technology[J]. Journal of Ningbo University (NSEE), 26(4): 17–23 (in Chinese with English abstract).
阮国良, 杨代勤, 王卫民, 2012. 草鱼、鳡和翘嘴鲌消化道组织的早期发育[J]. 水生生物学报, 36(6): 1164–1170. RUAN GUOLIANG, YANG DAIQIN, WANG WEIMIN, 2012. Ontogeny of the digestive tracts in Grass carp (), Yellowcheck carp () and Topmouth culter ()[J]. Acta Hydrobiologica Sinic, 36(6): 1164–1170.
石戈, 王健鑫, 刘雪珠, 等, 2007 褐菖鲉消化道的组织学和组织化学[J]. 水产学报, 31(3): 293–302. SHI GE, WANG JIANXIN, LIU XUEZHU, et al, 2007. Study on histology and histochemistry of digestive tract in[J]. Journal of Fisheries of China, 31(3): 293–302 (in Chinese with English abstract).
孙文静, 王晓艳, 祁鹏志, 等, 2018. 苯并[]芘(BaP)对褐菖鲉()肝CYP1A1酶活性、基因表达及蛋白表达的影响[J]. 海洋与湖沼, 49(4): 897–903. SUN WENJING, WANG XIAOYAN, QI PENGZHI, et al, 2018. Effects of benzo[]pyrene on erod activity, mRNA expression, and protein expression of cyp1a1 in the liver of[J]. Oceanologia et Limnologia Sinica, 49(4): 897–903 (in Chinese with English abstract).
王永翠, 李加儿, 区又君, 等, 2012. 黄鳍鲷仔、稚、幼鱼消化道形态组织学观察[J]. 南方农业学报, 43(8): 1212–1217. WANG YONGCUI, LI JIAER, OU YOUJUN, et al, 2012. Histomorphology observation on digestive tract of larva, juvenile and young Yellowfin black porgy ()[J]. Journal of Southern Agriculture, 43(8): 1212–1217 (in Chinese with English abstract).
谢木娇, 区又君, 李加儿, 等, 2017. 四指马鲅()消化系统胚后发育组织学观察[J]. 渔业科学进展, 38(2): 50–58. XIE MUJIAO, OU YOUJUN, LI JIAER, et al, 2017. Histological observation of the post-embryonic development of digestive tract of[J]. Progress in Fishery Sciences, 38(2): 50–58 (in Chinese with English abstract).
徐革锋, 陈侠君, 杜佳, 等, 2009. 鱼类消化系统的结构、功能及消化酶的分布与特性[J]. 水产学杂志, 22(4): 49–55. XU GEFENG, CHEN XIAJUN, DU JIA, et al, 2009. Fish digestive system: it’s structure, function and the distributions and characteristics of digestive enzymes[J]. Chinese Journal Fisheries, 22(4): 49–55 (in Chinese with English abstract).
徐晓津, 王军, 谢仰杰, 等, 2010. 大黄鱼消化系统胚后发育的组织学研究[J]. 大连水产学院学报, 25(2): 107–112. XU XIAOJIN, WANG JUN, XIE YANGJIE, et al, 2010. Post-embryonic histological development in digestive system of Large yellow croaker[J]. Journal of Dalian Fisheries University, 25(2): 107–112 (in Chinese with English abstract).
姚俊杰, 梁正其, 冯亚楠, 等, 2013. 普安银鲫消化系统胚后发育的组织学观察[J]. 贵州农业科学, 41(11): 152–155. YAO JUNJIE, LIANG ZHENGQI, FENG YA’NAN, et al, 2013. Histological studies on post-embryonic development for digestive system of[J]. Guizhou Agricultural Sciences, 41(11): 152–155 (in Chinese with English abstract).
张永泉, 刘奕, 尹家胜, 等, 2010. 哲罗鱼()消化系统胚后发育的形态与组织学的研究[J]. 海洋与湖沼, 41(3): 422–428. ZHANG YONGQUAN, LIU YI, YIN JIASHENG, et al, 2010. Morphology and histology of post-embryonic digestive system of[J]. Oceanologia et Limnologia Sinica, 41(3): 422–428 (in Chinese with English abstract).
ANDERSON S A, SALINAS I, WALKER S P, et al, 2012. Early development of New Zealand hapukueggs and larvae[J]. Journal of Fish Biology, 80(3): 555–571.
BOULHIC M, GABAUDAN J, 1992. Histological study of the organogenesis of the digestive system and swim bladder of the Dover sole,(Linnaeus 1758)[J]. Aquaculture, 102(4): 373–396.
DÍAZ A O, GARCÍA A M, GOLDEMBERG A L, 2008. Glycoconjugates in the mucosa of the digestive tract of: a histochemical study[J]. Acta Histochemica, 110(1): 76–85.
FACCIOLI C K, CHEDID R A, MORI R H, et al, 2016. Organogenesis of the digestive system in Neotropical carnivorous freshwater catfish(Siluriformes: Pimelodidae)[J]. Aquaculture, 451: 205–212.
GISBERT E, PIEDRAHITA R H, CONKLIN D E, 2004. Ontogenetic development of the digestive system in California halibut () with notes on feeding practices[J]. Aquaculture, 232(1–4): 455–470.
MICALE V, LEVANTI M B, GERMANÀ A, et al, 2010. Ontogeny and distribution of cholecystokinin-immuno reactive cells in the digestive tract of sharpsnout sea bream,(Cetti, 1777), during larval development[J]. General and Comparative Endocrinology, 169(1): 23–27.
MURAKAMI T, MORITA Y, 2010. Morphology and distribution of the projection neurons in the cerebellum in a teleost,[J]. Journal of Comparative Neurology, 256(4): 607–623.
MURAKAMI T, MORITA Y, ITO H, 1983. Extrinsic and intrinsic fiber connections of the telencephalon in a teleost,[J]. Journal of Comparative Neurology, 216(2): 115–131.
MURRAY H M, WRIGHT G M, GOFF G P, 1996. A comparative histological and histochemical study of the post‐gastric alimentary canal from three species of pleuronectid, the Atlantic halibut, the yellowtail flounder and the winter flounder[J]. Journal of Fish Biology, 48(2): 187–206.
PEARSE A G E, 1983. Histochemistry, theoretical and applied[M]. 4th ed. London: Churchill Livingstone: 96–101.
SARASQUETE M C, POLO A, YÚFERA M, 1995. Histology and histochemistry of the development of the digestive system of larval gilthead seabream,L.[J]. Aquaculture, 130(1): 79–92.
SOLOVYEV M M, CAMPOVERDE C, ÖZTÜRK S, et al, 2016. Morphological and functional description of the development of the digestive system in meagre (): an integrative approach[J]. Aquaculture, 464: 381–391.
TANAKA M, 1971. Studies on the structure and function of the digestive system in teleost larvae. III. Development of the digestive system during postlarval stage[J]. Japanese Journal of Ichthyology, 18(4): 164–174.
ZHENG RONGHUI, CHEN HUANBIN, BO JUN, et al, 2016. Joint effects of crude oil and heavy metals on the gill filament EROD activity of marbled rockfish[J]. Ecotoxicology and Environmental Safety, 132: 116–122.
Histological studies on development of the digestive system in larval and juvenile
YANG Jiazhe1, QI Chuang1, XU Shanliang2
1. School of Marine Sciences, Ningbo University, Ningbo 315211, China; 2. Key Laboratory of Applied Marine Biotechnology, Ningbo University, Ningbo 315211, China
as a marine economical teleost, research on the development of thelarvae and juveniles of this species become important. In this study, the developmental characteristics of digestive system in.from 0 to 50 days post-parturition (dpp) were described by the method of tissue sectioning technique, HE (hematoxylin-eosin) staining and histochemical staining. The results showed that the initial digestive system appeared to have a differentiated buccopharyngeal cavity after parturition. At 2 dpp, the larvae began feeding, and the digestive tube opened to the outside initially, and the larvae entered the endo-exotrophic period. The pyloric caeca appeared at 3-4 dpp, while 5~6 mucosal folds appeared in esophagus, and five mucosal folds appeared in stomach and hepatic cell mass grew. At 5-6 dpp, the yolk sac was completely absorbed, meaning the larvae began getting into exotrophic period Ⅰ (without a functional stomach). Meanwhile, goblet cells were observed in esophagus, and intestine can be divided into promesenteron and hindgut. At 10-14 dpp, circular layers of striated muscle were obviously observed and 7-12 mucosal folds appeared in esophagus; stomach had a basic structure, brush border became clear and there were a few hepatic sinusoids in a bigger liver, which was good for the digestion and absorption of exogenous food. At 28-30 dpp, the gastric glands and gastric pits appeared, which was the signs of the juvenile stage. With the increase of the number of islet cells and zymogen granules, the digestion was greatly improved. At 47-50 dpp, the digestive system gradually improved as a functional and structural one, which resembled that of adults. These results showed that the development of the digestive system in.started early like other ovoviviparous teleosts and was relative with the improving function of its digestion.
; larva and juvenile; digestive system; histology
2018-07-17;
2018-11-01. Editor: YIN Bo
Zhejiang “13th Five-Year” Advantage Professional Construction Project (Aquaculture Major of Ningbo University); Public Technology Application Research Projects of Zhejiang Province (2017C32015)
P735.541
A
1009-5470(2019)02-0058-09
10.11978/2018071
2018-07-17;
2018-11-01。殷波编辑
浙江省“十三五”优势专业建设项目(宁波大学水产养殖专业); 浙江省公益技术应用研究项目(2017C32015)
杨佳喆(1993—), 汉族, 男, 浙江省嘉兴市人, 研究生, 研究方向是海水鱼类健康养殖。Email: 1611091837@nbu.edu.cn
徐善良(1962—), 教授。E-mail: xushanliang@nbu.edu.cn
XU Shanliang. Email: xushanliang@nbu.edu.cn
