7个橡胶树品系树皮结构与农艺性状相关性研究
2019-04-08胡欣欣李维国王祥军张源源高新生
胡欣欣,李维国,王祥军,张源源,黄 肖,高新生
(1.中国热带农业科学院 橡胶研究所,海南 儋州 571737;2.海南大学热带农林学院,海南 海口 570228;3.国家橡胶树育种中心,海南 儋州 571737)
巴西橡胶树Hevea brasiliensis是原产南美亚马逊流域的热带高大乔木, 其主要经济产品是一种含橡胶的细胞质,即切割树皮中的次生乳管合成和积累的胶乳,其所提炼的天然橡胶是重要的工业原料和战略资源[1-2]。作为一种育种周期较长的经济作物,胶乳产量和茎围生长一直是橡胶树最重要的农艺性状。当前,聚焦以橡胶产量和木材产量双重目标的胶木兼优品种的选育已成为世界橡胶树品种选育的一个最重要的方向。
早在1923年,Bobilioff[3]首次指出了橡胶树的胶乳的产量与树皮的解剖结构的特性有关;后续多人研究证明橡胶树树皮结构中次生乳管列数目与天然橡胶产量高度相关[4-5];同时,环境因子、割胶、机械损伤、茉莉酸类物质等对橡胶树次生乳管也有很大影响[6-11]。成龄橡胶树树皮中分布大量的石细胞,其细胞数量多少,出现早晚是橡胶树品系特征之一。在同一橡胶树的不同部位的树皮中石细胞的形成也有明显的差异[12-13],同时石细胞也受环境因子、割胶、机械损伤、茉莉酸类物质的影响。近年来也有人研究,证实了橡胶树产量与茎围大小呈显著正相关[14]。基于橡胶树大田系统鉴定的传统品种评价方法所需周期较长,通过以农艺性状密切相关的树皮乳管分化能力辅助评价品系特性一直是橡胶树的一个重要研究方向。
目前有关橡胶树品系的次生乳管分化相关研究中,绝大多数只统计到石细胞团以内的乳管列数,本研究在参考郝秉中等[16-17]人方法的基础上,以我国自主选育的热试662、热试419、热试9359、热试647、热试451等5个无性系为试材,观测了其树皮乳管列数、厚度等次生乳管分化能力的相关参数,分析了上述参数与胶乳产量、茎围等主要农艺性状的相关性,旨在为以树皮乳管分化指标辅助评价主要农艺性状提供参考,同时也为参试品系配套割制等配套栽培技术的开发提供依据。
1 材料与方法
1.1 材 料
实验地点为中国热带农业科学院试验场二队的高级系比区,定植时间为2006年,开割时间为2013年,参试的7个橡胶树品系分别为热试662、热试419、热试647、热试9359和热试451,以RRIM600和PR107作为对照。开割时离地1.3 m处割胶,采用s/2 d3的非刺激割制。
参试材料的遗传关系如图1 所示,其中4个供试品系的亲本比较相近,热试662与热试662为同一亲本,热试419与热试451为同一亲本。从上述品系中各选10棵健康植株,定株分析其乳管分化能力及农艺性状的相关性。

图1 7个巴西橡胶树无性系的亲本谱Fig.1 Parental spectrum of seven clones of Hevea brasiliensis
1.2 方 法
1.2.1 树皮样品采集
试验材料在2017年开割后的6月采集树皮,方法为在割线上方10 cm左右用直径1.5 cm的打孔器采集。
1.2.2 树皮结构观察
以直径1.5 cm 打孔器在树干距离地面 150 cm上垂线位置取树皮样本,放入80%酒精固定,并经含碘和溴的冰醋酸溶液处理,然后用酒精梯度脱水,正丁醇透明,最后石蜡包埋。最后用旋转切片机进行制片,每一样品取3个玻片,在显微镜下观察树皮结构[18]。
1.2.3 观测参数
每株树的树皮标本做一张切片,每张切片统计 5 个不同视野。统计的参数如下:1)石细胞以内次生乳管列数目(NSLWS);2)石细胞以外的次生乳管列数目(NSLOS);3)树皮的次生乳管列数目(TC);4)石细胞以内的次生韧皮部厚度(mm)(TSLWS);5)石细胞以外的次生韧皮部的乳管列数的厚度(mm)(NTSLOS);6)石细胞以外的没有乳管列数的次生韧皮部的厚度(TSLOS);7)树皮的总厚度(TT);8)割线长度(cm)(SL);9)茎围(cm)(SG);10)原生皮的厚度(μm)(TV);11)再生皮厚度(μm)(TR);12)胶乳产量(mL)(LY);13)石细胞以内的次生乳管列数目与石细胞以内的次生韧皮部厚度的比值(NSLWS /TSLWS);14)石细胞以外的次生乳管列数目与石细胞以外的次生韧皮部厚度的比值(NSLOS/ NTSLOS);15)树皮中的次生乳管列数目与树皮总厚度的比值(TC/TT)。
1.2.4 胶乳产量
统计取样株上一年度即2016―2017年株次产胶量的均值作为该品系胶乳产量。
1.2.5 割线长度和茎围测量
在2017年7月底,使用测量尺于树干距地面1.5 m处测量。
1.2.6 树皮厚度测量
2017年7月底,以游标卡尺对割线垂直上方2 cm原生皮和上年度割面中间位置再生皮的厚度进行测量[18]。
1.2.7 统计方法
数据用SPSS V22 和excel V2010软件进行统计分析,采用 Duncan新复极差法对数据进行差异显著性检验。
2 结果与分析
2.1 各品系乳管分化参数的多重比较
从树皮的横切面看(图2),各品种石细胞以内的乳管列数目排列整齐和紧密。从表1可以得出,从石细胞以内的次生乳管列数目来看,石细胞以内的次生乳管列数目为8.56~19.89,其中PR107品种与热试662,热试9359,热试451等品种呈显著差异;从石细胞以外的次生乳管列数目来看,石细胞以外的次生乳管列数目为4.89~12.11,其中热试647品种与热试419,热试451,PR107,热试9359品种的呈显著差异;从树皮的次生乳管列数目来看,树皮结构次生乳管列数目为15.44~31.89,其中热试647,PR107,RRIM600品种与其他4个品种呈显著差异;从石细胞以内次生韧皮部的厚度来看,石细胞以内的次生韧皮部厚度为0.97~1.96 mm,其中PR107与其他6个品种呈显著差异;从石细胞以外的次生韧皮部的乳管列数目的厚度来看,石细胞以外的次生韧皮部的乳管列数目的厚度为1.98~3.11 mm,其中热试647品种与RRIM600品种无显著差异;从石细胞以外的没有乳管列数目的次生韧皮部的厚度来看,石细胞以外的没有乳管列数目的次生韧皮部的厚度为1.75~2.63 mm,其中热试451品种与热试662品种呈显著差异;从树皮的总厚度来看,树皮总厚度为5.69~7.44 mm,其中热试647品种与热试662,热试9359,热试451,RRIM600品种呈显著差异;从石细胞以内的次生乳管列数目与石细胞以内的次生韧皮部厚度的比值来看,石细胞以内的次生乳管列数目与石细胞以内的次生韧皮部厚度的比值为6.83~11.13,其中热试647和RRIM600品种与其他4个品种呈显著差异;从石细胞以外的次生乳管列数目与石细胞以外的次生韧皮部厚度的比值来看,石细胞以外的次生乳管列数目与石细胞以外的次生韧皮部厚度的比值为2.44~4.30,各品种之间无显著差异;从树皮的次生乳管列数目与树皮的总厚度的比值来看,树皮的次生乳管列数目与树皮的总厚度的比值为2.71~4.92,其中热试647品种与PR107和RRIM600品种无显著差异。
具体从品种来看,热试451和热试419品种的石细胞以内的次生乳管列数目,树皮的次生乳管列数目,石细胞以内的次生乳管列数目与石细胞以内的次生韧皮部厚度的比值,树皮的次生乳管列数与树皮的总厚度的比值等参数均较高,石细胞以外的没有乳管列数目的次生韧皮部的厚度和树皮的总厚度与其他品种相比相对较低。
2.2 各品系农艺性状参数的多重比较
从表2可以看出,各品系的茎围,原生皮厚度,再生皮厚度数值较为相近。从胶乳产量来看,胶乳产量为68.77~320 ml,其中热试419品种与其他6个品种呈显著差异;从割线长度来看,割线长度为32.94~38.22 cm,各品种之间无显著差异;从茎围来看,茎围为58.78~ 68 cm,各品种之间无显著差异;从原生皮的厚度来看,原生皮的厚度为8.18~8.87 μm,各品种间无显著差异;从再生皮的厚度来看,再生皮的厚度为5.85~7.77 μm,其中热试9359和RRIM600品种与热试647和热试419品种呈显著差异。
具体从品种来看,热试419品种的胶乳产量,割线长度,茎围最高,而原生皮的厚度和再生皮的厚度相对其他6个品种相对较低;热试9359和RRIM600品种的割线长度,原生皮的厚度,再生皮的厚度均较高,具有一定的产胶潜力。

图2 不同品系树皮的横切面Fig.2 Cross sections of barks of different clones
2.3 橡胶树树皮乳管分化参数的主成分分析
主成分分析可对多变量系统进行有效综合和简化,将多个指标转变为少数独立的综合指标,避免指标间的信息重叠。如上述10个树皮乳管分化参数去评价其乳管分化能力,效率低,且操作性差。因此,对上述参数进行主成分分析,取大于1的特征值,共获得4个主成分(表3),即4个综合指标。第 1 成分的贡献率为46.911%,第2 成分的贡献率为17.305%,第 3 成分的贡献率为14.057%,第 4 个成分的贡献率为12.970%,前 4个成分的累计贡献率已经达到91.242%,可见前4个成分足以说明该数据的变化趋势完全符合主成分分析的要求,故取前 4 个主成分作为数据分析的有效成分。

表1 巴西橡胶树品系乳管分化参数的方差分析†Table1 Variance analysis on milk duct differentiation parameters of Brazilian rubber tree clone stains

表2 各品系农艺性状的方差分析†Table2 Variance analysis on agronomic traits of different clone stains
由表4 可以看出,第1主成分贡献最大的是树皮的次生乳管列数目,树皮的次生乳管列数目与树皮的总厚度比值,石细胞以内次生乳管列数目,石细胞以外的次生乳管列数目,负荷量分别为0.989、0.911、0.849、0.721;第2主成分贡献最大的是石细胞以内的次生韧皮部厚度,负荷量为0.777;第3个主成分贡献最大的是石细胞以外的次生韧皮部的乳管列数目的厚度,负荷量为0.919;第4个主成分贡献最大的是石细胞以外的没有乳管列数目的次生韧皮部的厚度,负荷量为0.622。主要提取第1和2主成分,所以可以认为树皮结构的主要参数是树皮结构的次生乳管列数,树皮的次生乳管列数目与树皮的总厚度比值,石细胞以内次生乳管列数目,石细胞以外的次生乳管列数目,石细胞以内的次生韧皮部厚度。
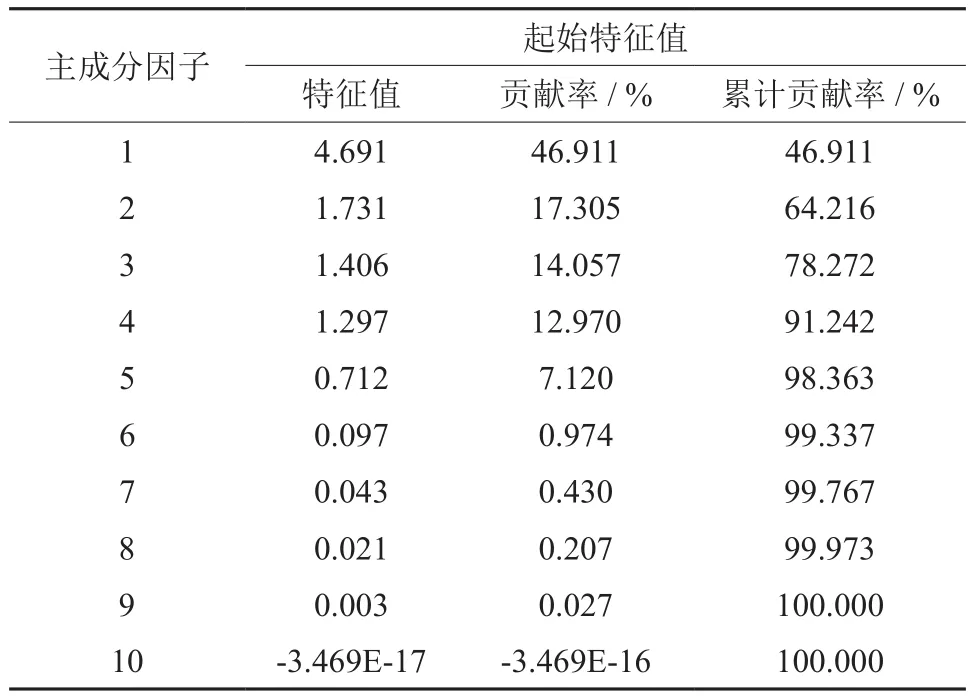
表3 不同性状的特征值Table3 Eigenvalues of different stain’s traits

表4 主成分载荷矩阵Table4 Principal component loading matrix
用x1,x2,x3,x4,x5,x6,x7分 别 表示 原先的7个品系,7个品系分别是热试662、热试419、热试9359、热试647、热试451,PR107、RRIM600。而用Y1,Y2,Y3,Y4表示新的主成分,那么这4个主成分(表6 )为:
Y1=0.028 889x1-0.181 110x2+0.057 778x3+0.131 111
x4+0.132 222x5-0.114 44x6-0.07x7;
Y2=0.181 111x1+0.072 222x2-0.033 33x3-0.144 440
x4-0.017 780x5-0.235 56x6+0.191 111x7;
Y3=0.067 778x1-0.092 22x2+0.207 778x3-0.055 562
x4-0.126 67x5+0.01x6+0.031111x7;
Y4=0.285 556x1-0.161 11x2-0.086 67x1-0.166 67
x4-0.098 89x5-0.163 333x6-0.018 89x7。
由表5得知,第1主成分得分最高的品系为热试451,第2主成分得分最高的品系为RRIM600,第3主成分得分最高的品系为热试9359,第4主成分得分最高的品系为热试662。
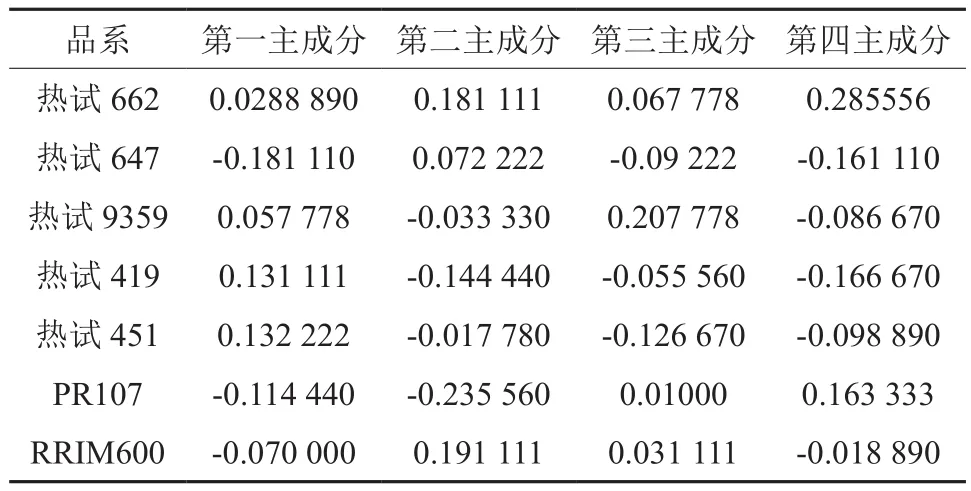
表5 主成分得分Table5 Principal component scores
2.4 各品系树皮乳管分化主成分与农艺性状的相关性
从表6可以看出,树皮的次生乳管列数目与石细胞以内的次生乳管列数目及石细胞以外的次生乳管列数目呈正相关;树皮中的次生乳管列数目与树皮总厚度的比值,与石细胞以内的次生乳管列数目呈正相关,且与石细胞以外次生乳管列数目和次生乳管的总列数目呈极显著正相关;石细胞以内的次生韧皮部厚度与茎围、再生皮的厚度呈正相关;胶乳产量与石细胞以内的次生乳管列数目,树皮中的次生乳管列数目与树皮总厚度的比值呈正相关。
3 结论与讨论
橡胶树的乳管系统是合成和贮藏橡胶胶乳的组织,树皮内的次生乳管是产胶的基础。在天然橡胶生产中,树皮的乳管列数目与胶乳产量密切相关[18]。己研究过的相关参数中,在幼苗中初生乳管的列数目与成龄树的产量有一定相关性,可以作为橡胶树育种早期选择的重要指标[18-19]。次生乳管列数目一般统计由形成层到石细胞出现的位置的乳管列数目[15]。卢世香等[15,20]认为,最内层石细胞团以内的次生韧皮部中乳管列数目与最内层石细胞团以内的次生韧皮部厚度的比值与胶乳产量成正相关。笔者在借鉴此方法的基础上,统计了石细胞以内次生乳管列数目,次生乳管列数目,树皮的总列数目与总厚度比值等10个参数,观测不同品系的乳管分化能力。其中,从石细胞以内的次生乳管列数目来看,石细胞以内的次生乳管列数目为8.56~19.89,其中PR107品种与热试662、热试9359、热试451等品种呈显著差异;从树皮的次生乳管列数目来看,树皮结构次生乳管列数目为15.44~31.89,其中热试647、PR107、RRIM600品种与其他4个品种呈显著差异;从树皮的次生乳管列数目与树皮的总厚度的比值来看,树皮的次生乳管列数目与树皮的总厚度的比值为2.71~4.92,其中热试647品种与PR107和RRIM600品种无显著差异。
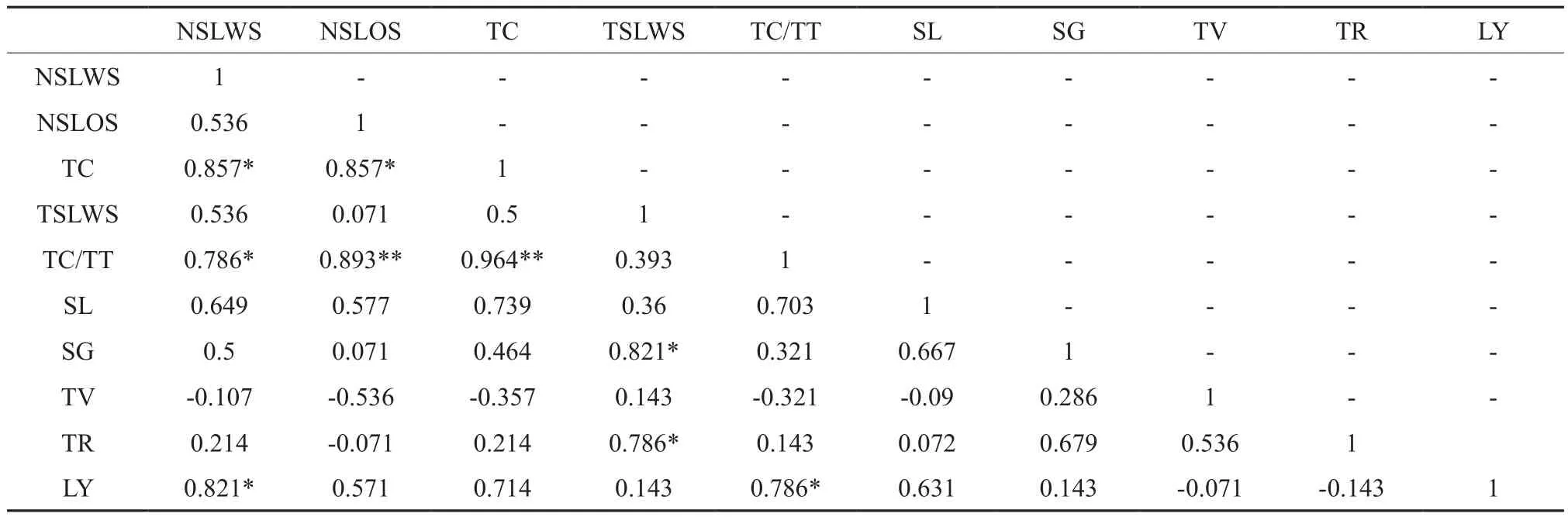
表6 树皮结构的主要因子与农艺性状的相关系数†Table6 Correlation coefficients of main factors and agronomic traits of tree bark structure
鉴于乳管的列数目与产胶量密切相关,乳管分化能力强的特异性种质能为其合成橡胶和贮存胶乳提供有利的条件,因此建立起橡胶树乳管分化能力的简便评价方法并用于特异性种质的筛选,对于橡胶树的早期预测及无性系系统鉴定具有重要的意义。针对橡胶树乳管分化能力评价参数较多,主效因子不明确的问题,本研究采用了主成分分析方法研究了不同品系间树皮乳管系统相关参数,对多变量系统进行有效综合和简化,将多个指标转变为少数独立的综合指标,避免指标间的信息重叠,继而找出引起树皮乳管分化的主要参数,结果表明,其主要参数分别为树皮的次生乳管列数,树皮的次生乳管列数目与树皮的总厚度比值,石细胞以内次生乳管列数目,石细胞以外的次生乳管列数目,石细胞以内的次生韧皮部厚度。通过主成分分析找出上述参数中的主效因子,并分析乳管分化各因子与品种农艺性状间的关系,对该乳管分化能力的评价方法进行了进一步拓展与完善,也为该方法应用于橡胶树品种选择提供了参考。结果表明,树皮中的次生乳管列数目与树皮总厚度的比值,与石细胞以内的次生乳管列数目呈正相关,且与石细胞以外次生乳管列数目和次生乳管的总列数目呈极显著正相关;石细胞以内的次生韧皮部厚度与茎围,再生皮的厚度呈正相关;胶乳产量与石细胞以内的次生乳管列数目,树皮中的次生乳管列数目与树皮总厚度的比值呈正相关。比如,热试419品系的石细胞以内的乳管列数较多,树皮中次生乳管列数目与树皮总厚度比值较大,胶乳产量最高;热试9359和RRIM600品系的石细胞以内的次生次生韧皮部的厚度较厚,茎围就较大,再生皮的厚度就较厚,产胶潜力较大。上述研究,进一步明确了乳管分化指标与重要农艺性状之间的关系,也对不同品系乳管分化能力有了系统了解。
结合参试材料的遗传背景来看,本研究7个参试无性系中,由于次生乳管的特征表现分化,其中热试419与热试451属于同一亲本组合,品种间胶乳产量有显著差异,其他参数均无显著差异;而热试662和热试647属于同一亲本,品种之间的石细胞以内的次生乳管列数目,树皮的次生乳管列数目,石细胞以外的次生韧皮部的乳管列数目的厚度,树皮的总厚度,石细胞以内的次生乳管列数目与石细胞以内的次生韧皮部的厚度比值,树皮的次生乳管列数目与树皮的总厚度的比值呈显著差异。RRIM600、PR107和热试9359品种之间亲缘关系不相近并且与以上4个品种亲缘关系较远。在农艺性状方面,同组合无性系虽然在茎围、树皮生长速度方面无统计学差异,但在产量方面分化更为明显。笔者认为,树皮结构作为无性系的遗传特性,品系间树皮结构的相似性应与其遗传背景相似性有很大关系,本研究中,两个相同组合子代的无性系在乳管相关参数表现分化,一方面在于橡胶树基因型高度杂合,作为同组合子代的F1代会产生基因分离,其基因型和表型均会有很大差异,一方面也在于样本量太少。而在农艺性状方面,尤其在产量性状方面,除受基因型控制外,也受其他重要因素影响,如当前橡胶树种苗繁育均属无性繁殖,砧木一般采用GT1的F1代种子苗,因此砧木基因型各不相同,而砧木和上部接穗部分会产生较强的互作效应,导致了即使在同一品种内,株间产量也存在较大的变异,这均导致了品系间遗传背景相似而产量差别显著。结果表明,通过主成分分析和相关分析可以得出,在7个品系中,石细胞以内的次生韧皮部厚度与茎围,再生皮的厚度呈正相关;胶乳产量与石细胞以内的次生乳管列数目,树皮中的次生乳管列数目与树皮总厚度的比值呈正相关。其中,热试419品系树皮结构的次生乳管列数目与树皮结构的总厚度的比值明显高于其他品系,次生乳管的总列数最多,乳管分化能力较强,且胶乳产量最高;热试9359和 RRIM600品系的次生乳管的分化能力较强,产胶潜力大。该研究对乳管分化指标与重要农艺性状之间的关系起到积极辅助作用,但具体评价橡胶树胶乳产量还需考虑很多因素,如干胶含量,胶乳生理参数,内源激素等。
