Macroecological Patterns of Climatic Niche Breadth Variation in Lacertid Lizards
2019-03-27MengchaoFANGXiamingZHUYuDULinZHANGandLonghuiLIN
Mengchao FANG, Xiaming ZHU, Yu DU, Lin ZHANG and Longhui LIN*
1 Hangzhou Key Lab for Ecosystem Protection and Restoration, College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 310036, China
2 Hainan Key Lab for Herpetological Research, College of Fisheries and Life Science, Hainan Tropical Ocean University, Sanya 572022, China
Abstract Measuring climatic niche position and breadth may help to determine where species can occur over space and time. Using GIS-based and phylogenetic comparative methods, we investigated global patterns of variation in climatic niche breadth in lacertid lizards to test the following three hypotheses about climatic niche widths. First, does a species’temperature or precipitation niche breadth relate to its temperature or precipitation niche position (the mean value of annual mean temperature or annual precipitation across sampled localities in the range of each species)? Second, are there trade-offs between a species’ temperature niche breadth and precipitation niche breadth? Third, does a species’temperature or precipitation niche breadth relate to altitude or latitude? We expect that: (1) species distributed in cold regions are specialized for low-temperature environments (i.e. narrow niche breadth center around low temperatures);(2) a negative relationship between species niche breadth on temperature and precipitation axes according to the tradeoff hypothesis (i.e. species that tolerate a broad range of precipitation regimes cannot also tolerate a broad range of temperatures); (3) precipitation niche breadth decreases with altitude or latitude, whereas temperature climatic niche breadth increases with altitude or latitude. Based on the analytical results we found that: (1) temperature niche breadth and position are negatively related, while precipitation niche breadth and position are positively related; (2) there is no trade-off between temperature and precipitation niche breadths; and (3) temperature niche breadth and latitude/altitude are positively related, but precipitation niche breadth and latitude/altitude are not significantly related. Our results show many similarities with previous studies on climatic niche widths reported for amphibians and lizards, which provide further evidence that such macroecological patterns of variation in climatic niche breadths may be widespread.
Keywords Macroecological pattern, climatic niche breadth, climatic niche position, Lacertidae
1. Introduction
Climatic niche breadth, the size of the range of temperature and precipitation conditions where a species occurs, may help to determine species occurrence patterns over space and time. In addition, the climatic niche breadth is vital concept to address many lines of inquiries such as: species persistence and diversification, species responses to anthropogenic climate change, and the colonization of invasive species (Bonetti and Wiens, 2014;
Dellinger et al., 2016; Gómez-Rodríguez et al., 2015;Soberón, 2007; Title and Burns, 2015; Velasco et al.,2016). For example, a lineage adapting more quickly to new climatic niches may have a decreased rate of extinction in a changing environment (Holt, 2009; Kozak and Wiens, 2010; Title and Burns, 2015), and species with narrow climatic niche breadths tend to have small distributional ranges (Li et al., 2016; Velasco et al., 2016).Macroecological patterns of climatic niche breadths seem to be similar in most amphibians, some lizards and some snakes (Bonetti and Wiens, 2014; Li et al., 2016; Lin et al., 2019; Quintero and Wiens, 2013; Title and Burns,2015; Wiens et al., 2013). For example, most amphibians(Bonetti and Wiens, 2014), phrynosomatid lizards(Quintero and Wiens, 2013; Wiens et al., 2013), varanid lizards (Lin and Wiens, 2017) and terrestrial elapid snakes(Lin et al., 2019) in humid environments have broader precipitation niche breadths than do closely related species in arid environments, and species occurring in colder environments have broader temperature niche breadths than do tropical species (Bonetti and Wiens,2014; Chejanovski and Wiens, 2014; Lin and Wiens,2017; Velasco et al., 2016; Wiens et al., 2013). In addition, these groups show a positive relationship between niche breadth in temperature and precipitation axes (i.e. temperature niche breadth increases with precipitation niche breadth; Bonetti and Wiens, 2014;Lin and Wiens, 2017; Lin et al., 2019). However, the generality of these macroecological patterns of climatic niche breadths need further evidence provided by different groups of organisms. Moreover, climate niche breadth may help drive patterns of species richness by affecting diversification rates. For example, niche divergence and rates of niche change explain more than half of variation in net diversification rates among mammal families(Castro-Insua et al., 2018). Specialist species had higher net diversification rates than generalist species in a recent study focusing on the relationship between temperature niche breadth and diversification for 2 340 species of amphibians, 4 563 species of mammals and 9 823 species of birds (Rolland and Salamin, 2016). Anole lizards with narrow niche breadths exhibited higher speciation rates than those with broader niche breadths (Velasco et al.,2016). Thus, measuring the climatic niche breadth helps to understand where a species can occur over space and time (Lavergne et al., 2010), the emergence of diversity patterns (Rolland and Salamin, 2016), dynamic species distributions, responses to climate change, and our ability to model future species distributions (Ralston et al.,2016).
In this study we focused on lacertid lizards, a widelydistributed family found in Africa, Europe and Asia.We tested three general hypotheses on variation in climatic niche width among species. First, does a species’temperature or precipitation niche breadth relate to its temperature or precipitation niche position (the mean value of annual mean temperature or annual precipitation across sampled localities in the range of each species)?For example, are species distributed in cold regions specialized for low-temperature environments (i.e. narrow niche breadth center around low temperatures)? Second,does a species’ temperature niche breadth positively or negatively relate to its precipitation niche breadth? (i.e.does a species with a broader temperature niche breadth have a broader or narrower precipitation niche breadth?)Third, does a species’ temperature or precipitation niche breadth relate to altitude or latitude? There are few studies on the relationship between climatic niche breadth and altitude, though Alahuhta et al. (2017) found a positive relationship between temperature niche breadth and lake altitude in aquatic macrophyte species in Sweden. For the relationship between niche breadth and latitude, Janzen(1967) contended that the climatic niche for temperature should be narrower in tropical species, while Vázquez and Stevens (2004) reported that the precipitation niche should be broader in tropical species.
2. Materials and Methods
We used the climatic data and phylogeny to test our hypotheses. We collected occurrence points for 137 species for lacertid lizards (~42% of all species described globally; Figure 1, Table S1) from HerpNet/VertNet(www.vertnet.org), Global Biodiversity Information Facility (www.gbif.org), and references involving the distribution of these lacertid lizards (references are listed in Table S1). All 137 species were included in the phylogeny of Pyron et al. (2013). We carefully checked occurrence points to ensure that sampled localities spanned each species’ full geographic range and that no localities were erroneously outside that range, according to the Reptile Database (www.reptile-database.org, Uetz and Hok, 2016). We extracted climatic variables for each occurrence point from the WorldClim database(www.worldclim.org, Hijmans et al., 2005) at ~1 km2resolution using DIVA-GIS 7.5.0. We obtained climatic data from a mean of 312.5 localities per species, with a range of 5-11071 localities (Table S1).
We chose latitude (LAT) and five climatic variables from the WorldClim database: altitude (ALT), annual mean temperature (AMT), maximum temperature of the warmest month (MTWM), minimum temperature of the coldest month (MTCM), and annual precipitation(AP) for this analysis. Mean values for AMT across a species’ range represent temperature niche positions of the species; mean values for AP represent precipitation niche positions. For latitude, we adopted the absolute value to calculate the mean latitude of each species across localities in its range when it was located in the Southern Hemisphere. We calculated temperature niche breadth and precipitation niche breadth for each species (Table S2) as follows:
Temperature niche breadth = maxMTWM - minMTCM

Figure 1 Point localities of the 137 species of terrestrial lacertid lizards involved in this study.
Precipitation niche breadth = maxAP- minAP where maxMTWM is the maximum value of MTWM across all sampled localities of a species, minMTCM the minimum value of MTCM, maxAP the maximum value of AP, minAP the minimum value of AP. We defined temperature niche position as the mean value of AMT across sampled localities in the range of each species, and precipitation niche position as the mean value of AP (Lin and Wiens, 2017).
We acquired the phylogeny of 137 species of lacertid lizards by trimming the phylogeny of Squamata which included 4 161 species of lizards and snakes (Pyron et al., 2013). We kept all the 137 species in the phylogenetic tree and excluded the species without locality data from the analysis by the program’s command in R 3.3.0 (R Development Core Team, 2015). We ran Levene’s test for homogeneity of variances on each pair of variables in Table 1 in SPSS 20 (Norušis, 2011). If variances were not equal across the datasets, we then performed Spearman’s test for rank correlation in SPSS 20. If variances were equal, we performed phylogenetic generalized least squares regression (PGLS) analyses in R 3.3.0 with the package Caper (Orme, 2013) to account for the statistical non-independence of species data due to phylogeny.We used the λ model for all PGLS analyses with branch lengths adjusted based on λ values estimated via maximum likelihood, and values of kappa and delta fi xed at one. We performed spatial autocorrelation analysis in DPS 14.10 (Tang and Zhang, 2013) to fi nd that none of Moran’s I values deviated significantly from 0 (all P >0.450), which indicated a lack of spatial autocorrelation.We performed resampling analyses to test if the climatic data for each species is influenced by species sampling following the methods described by Quintero and Wiens(2013). Random sampling was reiterated 1000 times for each species and climatic variable. Then we examined the relationship between samples size and the magnitude of the differences between the actual species value and the average simulated value.
3. Result
We found a marginally significant linear relationship for temperature niche breadth (R = 0.174, F1,122=3.798, P = 0.054) and a non-significant relationship for precipitation niche breadth (R = 0.098, F1,122= 1.179,P = 0.280) between samples size and the magnitude of the differences between the actual species value and the average simulated value. The P value obtained from Spearman’s test was consistent with that from PGLS in each case (Table 1). Globally, lacertid species in warmer climates with higher mean values for annual mean temperature generally have narrower temperature niche breadth (r2= 0.156, F1,135= 24.950, P < 0.0001;Figure 2A; Table 1). This suggests that lacertid species distributed in warmer regions are more specialized forhigh temperature environments. Lacertid species in more humid climates with higher mean values for mean annual precipitation generally have a broader precipitation niche breadth (r2= 0.189, F1,135= 31.490, P < 0.0001,Figure 2B; Table 1). This suggests that lacertid species distributed in more arid regions are more specialized for arid environments.
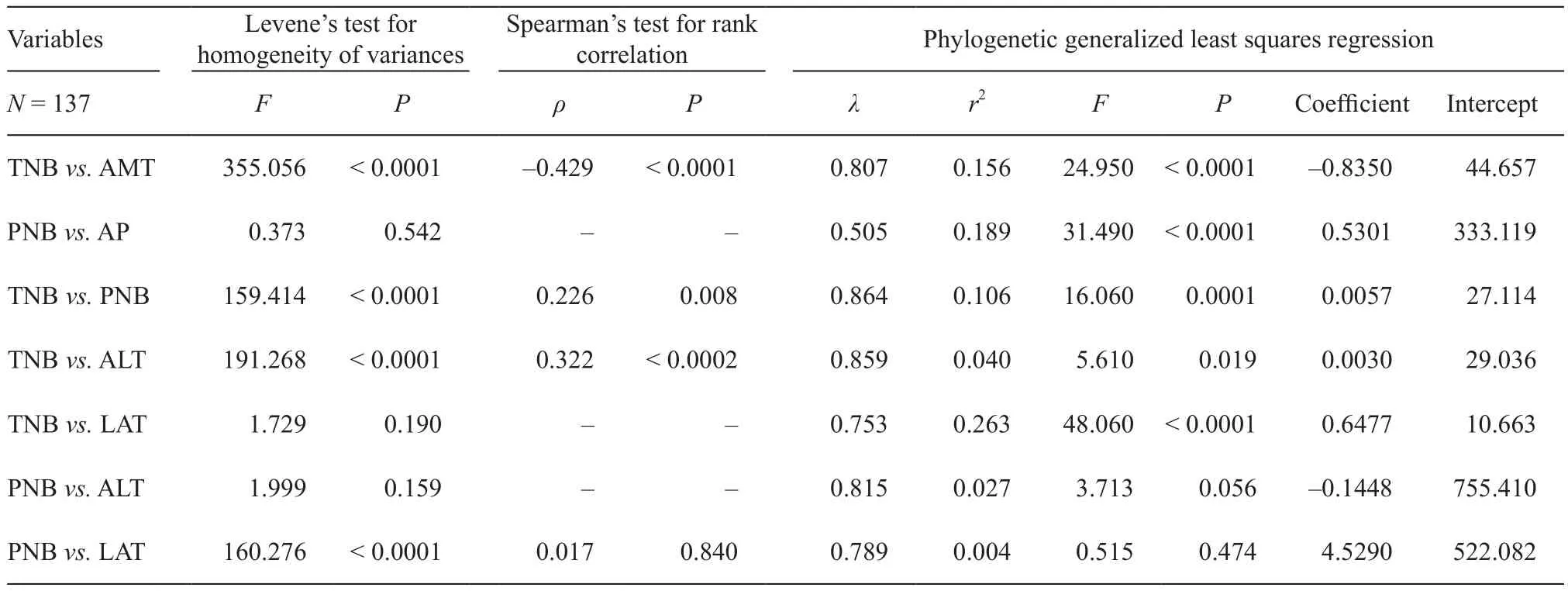
Table 1 Levene’s test for homogeneity of variances, Spearman’s test for rank correlation, and phylogenetic generalized least squares regression (PGLS) results of the relationship between temperature niche breadth (TNB) and annual mean temperature (AMT), between precipitation niche breadth (PNB) and annual precipitation (AP), between TNB and PNB, between TNB and altitude (ALT)/latitude (LAT),between PNB and ALT/LAT. N is the number of species. If variances were not equal across the datasets we then performed Spearman’s test,whereas the results of PGLS were also listed. The P value obtained from Spearman’s test was consistent with that from PGLS in each case.
Temperature niche breadth and precipitation niche breadth are positively related (r2= 0.106, F1,135= 16.060,P < 0.001; Figure 2C; Table 1), suggesting no tradeoff between temperature niche breadth and precipitation niche breadth. Lacertid species in high altitudes generally have a broader temperature niche breadth (r2= 0.040,F1,135= 5.610, P = 0.019; Figure 2D; Table 1), but there is not a significant relationship between altitude and precipitation niche breadth (r2= 0.027, F1,135= 3.713, P= 0.056; Table 1). This suggests that for lacertid species,precipitation niche breadth does not increase linearly with altitude (i.e. lacertid species living in higher altitudes do not necessarily have wider precipitation niche breadths).Lacertid species in higher latitudes generally have broader temperature niche breadths (r2= 0.263, F1,135=48.060, P < 0.001; Figure 2E; Table 1), but there is not a significant relationship between latitude and precipitation niche breadth (r2= 0.004, F1,135= 0.515, P = 0.474, Table 1). This suggests that lacertid species living in higher latitudes do not necessarily have higher precipitation niche breadths.
4. Discussion
We estimated climatic niche positions and breadth by measuring realized climatic niches, considering various abiotic (e.g. temperature, precipitation, air velocity and light intensity) and biotic factors (e.g. competition,predators and pathogens and mutualisms; see McGill et al. [2006] and Soberón [2007]). This approach allowed us to compare our results to those of previous studies on realized climatic niche breadths (Bonetti and Wiens,2014; Chejanovski and Wiens, 2014; Lin and Wiens,2017; Lin et al., 2019; Quintero and Wiens, 2013; Wiens et al., 2013).
We found that temperature niche breadth and position are negatively related. We suggest that at higher altitudes/latitudes with lower annual mean temperature, variation in temperature increases such that species have broader realized temperature niche breadth (Janzen, 1967; Lin and Wiens, 2017; Lin et al., 2019; Quintero and Wiens,2013; Vázquez and Stevens, 2004). On the other hand,precipitation niche breadth and position are positively related, as found in phrynosomatid lizards (Wiens et al.,2013), amphibians (Bonetti and Wiens, 2014), varanid lizards (Lin and Wiens, 2017) and terrestrial elapid snakes(Lin et al., 2019). This result corroborates the viewpoint that species in more mesic environments have a broader precipitation niche breadth (Bonetti and Wiens, 2014).

Figure 2 Relationships between temperature niche breadth and annual mean temperature (A), between precipitation niche breadth and annual precipitation (B), between temperature niche breadth and precipitation niche breadth (C), between temperature niche breadth and altitude (D), and between temperature niche breadth and latitude (E).
Moreover, we found that there is no trade-off between temperature and precipitation niche breadths, rejecting the hypothesis that there should be a trade-off in niche breadths on different niche axes (i.e. temperature and precipitation niche breadths negatively interrelated;Vázquez and Stevens, 2004). Conversely, we found a positive relationship between temperature niche breadth and precipitation niche breadth, as found in amphibians(Bonetti and Wiens, 2014; Chejanovski and Wiens,2014) and varanid lizards (Lin and Wiens, 2017). We suggest that species in more extreme environments might use similar mechanisms to cope with stress from both temperature and precipitation extremes, potentially leading to broad or narrow niche breadths on both axes(Bonetti and Wiens, 2014; Wiens et al., 2013).
Furthermore, temperature niche breadth and latitude/altitude are positively related; however, precipitation niche breadth and latitude/altitude are not significantly related. Temperature niche breadth increases with latitude in this study, as similar to hylid frogs, phrynosomatid lizards, and plethodontid salamanders (Quintero and Wiens, 2013). This finding was also consistent with Janzen’s hypothesis (1967; but see Vázquez and Stevens,2004). This result is also consistent with that of highaltitude tropical insects, which have broader temperature niche breadths because of summer-like conditions during the day and winter-like conditions at night every day of the year (Mani, 1968). Similarly, high-altitude tropical amphibians physiologically perform better over broader ranges of temperatures than do their low-altitude counterparts (Lüddecke and Sínchez, 2002). Quintero and Wiens (2013) found a negative relationship between precipitation niche breadth and latitude in phrynosomatid lizards (see also Vázquez and Stevens, 2004); however,they found that this relationship was not significant in plethodontid salamanders, similar to this study. Thus,the relationship between precipitation niche breadth and latitude/altitude needs further research in different groups of organisms such as aquatic vertebrates or invertebrates.
Exploring the relationships between climate niche breadth and climate niche position, latitude and altitude help to understand patterns of species diversification and species richness, dynamic species distributions, responses to climate change and our ability to model future species distributions. Future studies may focus on exploring the relationship between diversification rates and the climatic niches of species and clades. By measuring the niche change within clades such as rates of climatic niche divergence we could explain patterns of variation in diversification rates among clades.
Some caveats worth mentioning are the following: (1)For temperature niche breadth, there was a marginally significant linear relationship between sample size and the magnitude of the differences between the actual species value and the average simulated value. We should bear in mind that there is the limitation of our analyses neglecting the effect of the samples size even though we excluded species with fewer than 5 occurrence points. (2) Furthermore, R squared percentages in our results are extremely low, thus the data suggest a weak trend between the parameters. (3) Our results are based on patterns of climatic variation, and not directly on physiological tolerances. However, our goal is to explore these patterns of climatic distribution (e.g. the coldest/warmest temperatures where species occur), and not to estimate physiological parameters (e.g. active body temperatures) (see Quintero and Wiens, 2013). (4) We calculated climatic niche breadths using yearly extremes in precipitation. However, other aspects of those climatic variables (e.g. precipitation of the wettest quarter and precipitation of the driest quarter) were also likely to determine the range and distributions of climatic niche instead (see Quintero and Wiens, 2013).
Acknowledgements Financial support was provided by grants from Zhejiang Provincial Natural Science Foundation (LY17C030003), Natural Science Foundation of China (31270571), and Science and Technology Bureau of Sanya (2013YD08). We thank two anonymous referees for comments on this paper.
Appendix
Table S1 Summary of data on latitude, longitude, altitude, annual mean temperature (AMT), maximum temperature of the warmest month(MTWM), minimum temperature of the coldest month (MTCM), annual precipitation (AP) and references for the occurrence points per multi ple individuals of each species.
https://pan.baidu.com/s/1kIS-WLmYYG_UrsqqEywQqQ
Table S2 Summary of data on the number of geo-referenced localities per species (N), species niche breadths for temperature (TNB) and precipitation (PNB), species mean values for altitude, latitude, annual mean temperature (AMT) and annual precipitation (AP).
https://pan.baidu.com/s/1_7TL9vWWMPcWz4xEkPOn8Q
杂志排行

Asian Herpetological Research的其它文章
- Investigating the Effectiveness of Road-related Mitigation Measures under Semi-controlled Conditions: A Case Study on Asian Amphibians
- Embryonic Growth and Yolk Depletion during Incubation in the Chinese Skink, Plestiodon chinensis
- Microhabitat Segregation of Parapatric Frogs in the Qinling Mountains
- Female-biased Dispersal of the Emei Moustache Toad(Leptobrachium boringii) under Local Resource Competition
- A New Species of the Genus Trimeresurus from Southwest China(Squamata: Viperidae)
- Resurrection of the Genus Leptomantis, with Description of a New Genus to the Family Rhacophoridae (Amphibia: Anura)