曼氏无针乌贼染色体核型分析
2019-03-25高天翔王露萍迟长凤江丽华王世锋
高天翔,王露萍,迟长凤,江丽华,王世锋
(1.浙江海洋大学水产学院,浙江舟山 316022;2.国家海洋设施养殖工程技术研究中心,浙江舟山 316022;3.浙江海洋大学海洋科学与技术学院,浙江舟山 316022;4.海南大学海洋学院,海南海口 570228)
曼氏无针乌贼(Sepiella maindroni de Rochebrune)俗称乌贼、墨鱼、血墨。分布于渤海、黄海、东海、南海,为印度洋和西太平洋广分布种,其中心分布区在中国浙江近海和闽东海域。曼氏无针乌贼是东海重要经济种类,20 世纪50-70 年代曾是舟山渔场“四大海产”之一,但由于资源的不合理利用,以及生态环境的破坏,80 年代中后期,浙江沿海渔场曼氏无针乌贼产量急剧下降,已形不成渔汛,濒临灭绝[1]。21 世纪初,浙江海洋学院和宁波大学先后开展了曼氏无针乌贼人工繁育技术研究,并取得突破。2007-2009 年浙江海洋学院研究团队在浙江沿海的舟山东极海域、台州大陈海域、温州洞头海域开展小规模曼氏无针乌贼苗种(受精卵)探索性增殖放流研究[2]。近年来,福建省和江苏省也相继开展了曼氏无针乌贼的增殖放流工作。随着人工繁育技术的突破和人工增殖放流工作的开展,目前资源量得到了显著恢复,据相关报道资源量已恢复至2 000 t 的水平[3]。目前有关曼氏无针乌贼的研究工作主要集中在繁殖生物学[4-6]、增殖放流[1,7]及营养饲料[8-10]等方面。
染色体是生物发育、进化、遗传和变异的物质基础,研究染色体的行为、数目、核型和带型等对于了解生物的遗传组成、遗传变异规律、发育机制、物种起源和亲缘关系、进化地位、分类及种族关系等都具有重要的参考价值[11]。目前有关头足类染色体的研究国内外已开展较多研究:莱氏拟乌贼Sepioteuthis lessoniana[12]、长蛸Octopus minor[13]、金乌贼Sepia esculenta[14]、鹦鹉螺Nautilus macromphalus[15]等。但有关曼氏无针乌贼染色体核型分析的研究在国内外尚未见报道。本研究制备了曼氏无针乌贼染色体并进行了核型分析,以期为曼氏无针乌贼的遗传背景和资源保护提供种质特征方面的相关资料。
1 材料与方法
1.1 实验材料
曼氏无针乌贼亲体为舟山渔场的野生乌贼,选择体质健康、性成熟个体,在室内人工养殖,实验样品取自亲体产出的受精卵和不同发育阶段的胚胎。
1.2 实验方法
1.2.1 染色体标本的制备
参照王晓华等[14]的金乌贼染色体制备方法,并对其方法进行优化。选取红眼期曼氏无针乌贼胚胎,剥去卵膜,取出胚胎。将胚胎浸入0.04%的秋水仙素溶液,浸泡处理1 h,去净秋水仙素,用0.075 mol·L-1KCl低渗处理45 min,离心,移去低渗液,加入4℃预冷的卡诺氏国定液(甲醇:冰醋酸=3:1)固定,间隔15 min换固定液1 次,连续固定3 次,然后加入50%冰醋酸进行解离,解离时间为45 min。将解离好的样品热滴法滴片,空气干燥过夜。
1.2.2 染色体数目统计及核型分析
制备好的染色体玻片用10% Giemsa 染液染色20 min,蒸馏水冲洗,自然晾干后进行显微观察和拍照。选择分散良好、形态清晰、两条臂适当分开且数目完整的中期分裂相约40 个进行拍照。进行染色体计数、测量,并根据LEVAN A,et al[16]的分类标准进行染色体配对、分组,制作核型图。
2 结果
2.1 染色体数目
实验对40 个分散良好的中期分裂相进行染色体计数,曼氏无针乌贼二倍体染色体数为92 的约占所统计中期分裂相的91%,占绝对优势,由此可以判定曼氏无针乌贼的二倍体染色体数目为2n=92。
2.2 染色体核型
选取3 个分散良好,形态清晰的中期分裂相进行测量和统计分析,获得曼氏无针乌贼染色体的相对长度[染色体相对长度=(单条染色体长度×2/全部染色体总长度)×100]、臂比及其核型,如图1 和表1 所示。根据LEVAN,et al[16]提出的核型分类标准,可将曼氏无针乌贼染色体分为4 个类型,其中,m 型染色体28 对,sm 型12 对,st 型4 对,t 型3对,核型公式为2n=54m+24sm+8st+6t,臂数(NF)=170。

图1 曼氏无针乌贼染色体中期分裂相(A)及核型(B)Fig.1 Metaphase chromosomes (A) and karyotype (B) of S.maindroni

表1 曼氏无针乌贼染色体相对长度及臂比值(平均值±标准差)Tab.1 Relative length and ratio of chromosomes in S.maindroni
3 讨论
3.1 染色体的制备
提取到新陈代谢旺盛、分裂增生较快的组织是成功制备染色体的基本条件[17-18]。鱼类染色体的获得通常采用体内注射PHA 肾细胞培养法[18],但是由于头足类染色体数目多及形态大小适中的染色体中期分裂相较难获得等原因。据报道,用于头足类染色体的制备组织有3 种:(1)雄性的精细胞[19-20];(2)鳃心血液[17];(3)胚胎[12-13,21]。采用胚胎制作头足类染色体可以获得比较理想的结果[12]。本研究在制备曼氏无针乌贼染色体的过程中,分别选取了受精卵和不同发育时期的胚胎作为染色体制备材料。实验发现只有以红眼期的曼氏无针乌贼胚胎为材料制备的染色体中,染色体分散最好,中期分裂相对较多,且染色体的形态最为清晰。因此,本研究认为曼氏无针乌贼染色体标本的制备材料以红眼期胚胎最佳。
3.2 染色体核型与种质鉴定关系
染色体是遗传物质的主要载体,染色体核型研究是细胞遗传学的基础,染色体组型在很大程度上具有物种的特异性,特定的生物体具有特定数目、形态特征的染色体[22-23]。染色体数目和结构对于研究物种进化具有重要作用。目前国内外头足类染色体核型研究仅13 例(表2)。GAO Yuemian,et al[12]成功对5 种头足类染色体核型进行了描述,这5 种头足类染色体数目均为92 条,核型稍有差别。而在此之前的很多研究,由于未获得好的中期分裂相或者显微拍照质量等原因,仅进行了染色体数目描述。WANG Jinhai,et al[13]对3 种蛸类做了染色体核型分析,发现基于染色体核型进化距离和核型近似系数的头足类进化分析,与基于线粒体全序列的头足类进化树结果一致,可以作为传统分类学和分子系统分析的补充,有望成为常规系统发育的辅助手段。然而在鱼类研究中,由于鱼类核型基本都是端部着丝粒染色体,核型进化距离和近似系数法并不适用。已有的头足类染色体研究结果与基于线粒体细胞色素氧化酶亚基I 的建树结果相一致(图3),其中鹦鹉螺染色体数目为52 条,是头足类最为古老的物种,而八腕总目染色体数目均为60 条,十腕总目染色体数除阿拉伯乌贼染色体数目为68 条外,其余均为92 条。十腕总目染色体数目具有高度一致性,但是不同种类的核型不同,可作为鉴别不同种属的重要依据。目前,头足类分类最传统的方法是依据其形态特征,通过对个体的分节特征、度量特征和解剖学特征的测量和鉴定,依据这些特征的差异进行分类[24]。但是头足类在受外界刺激或死亡状态下体色容易改变;另外,大多数头足类的幼体阶段外部特征比较相似。这些问题都为仅仅依靠形态学的方法鉴别头足类种类带来困难。因此,在头足类种类鉴别过程中,无法通过形态特征进行区分时,可以考虑使用染色体核型的方法。染色体核型的多样化为头足类的系统分类和种质鉴定有着重要的辅助作用。

图3 基于线粒体细胞色素氧化酶亚基I(COI)的头足类进化树Fig.3 Phylogenetic relationships among the cephalopods based on mitochondrial COI gene sequence
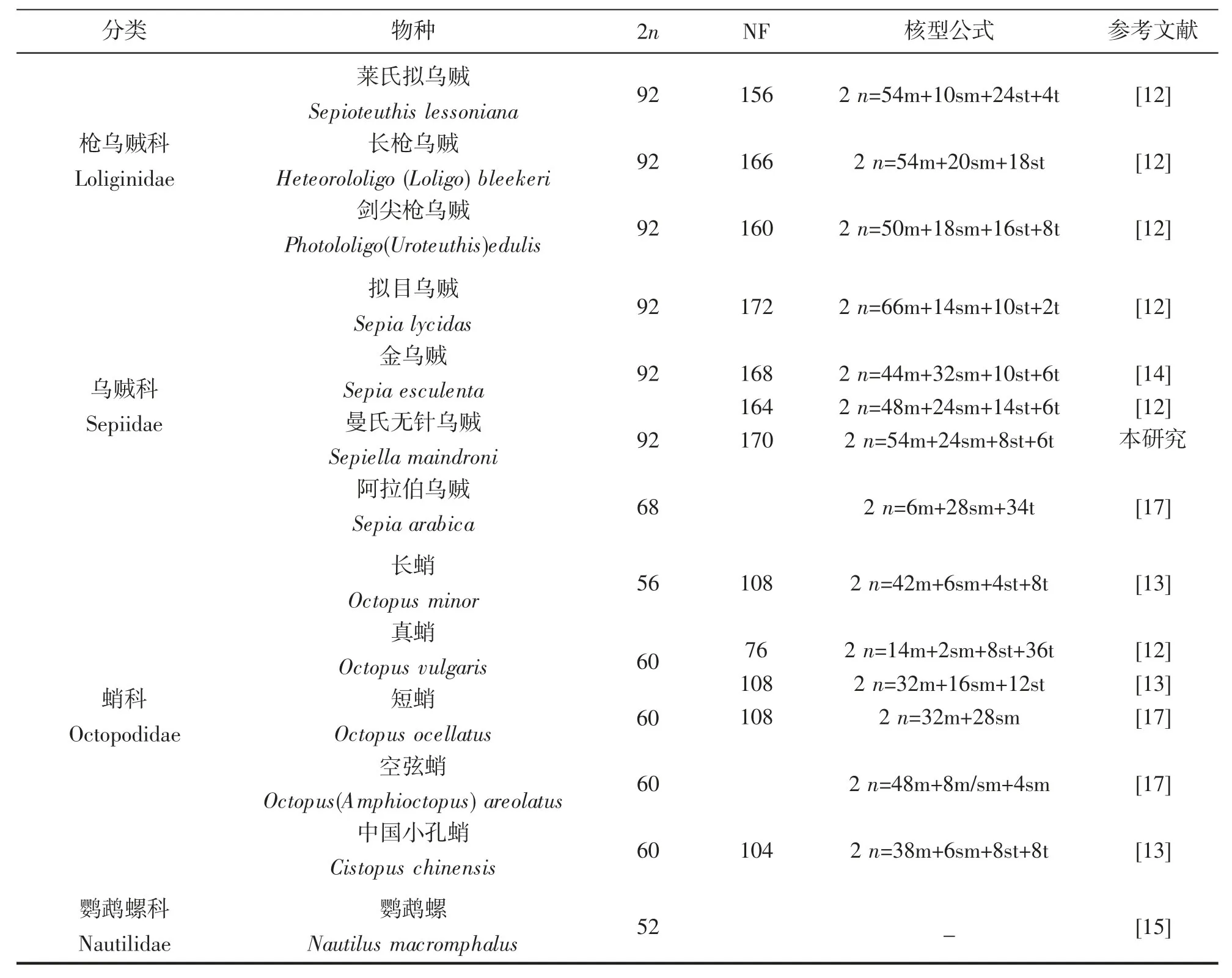
表2 部分已报道头足类染色体核型Tab.2 Karyotypes of several reported species of the cephalopods
为了更好研究头足类染色体遗传信息,在今后研究中应采用除染色体核型以外,开展带型研究、染色体核仁组织区及荧光原位杂交技术的综合分析。
