甜菜色素的生物学研究进展
2019-03-23徐夙侠黄青云林春松何恩铭
徐夙侠,黄青云,林春松,何恩铭
(福建省亚热带植物研究所/福建省亚热带植物生理与生化重点实验室,福建 厦门 361006)
甜菜色素是一类比较小众的色素,只在石竹目Caryophyllales的15个科的植物以及真菌类的毒蝇伞Amanita muscaria中发现[1—2],目前已鉴定出的甜菜色素种类有75种[3—4]。鉴于甜菜色素的分布特征,植物学家用它作为一种化学分类指标[5]。
1 甜菜色素的特性
甜菜色素是一类水溶性色素,与其他色素一样合成于细胞质中,储存于液泡[4,6],在植物体上能表现出红色、紫色、黄色、白色等丰富的色彩[7](图1: A, B)。目前,已知的甜菜色素都属于季胺型生物碱,分为以氨基酸为辅基类型的甜菜黄素(Betaxanthin)和糖苷化类型的甜菜红素(Betacyanin)两类[4,8](图1: C,D)。甜菜红素和甜菜黄素的吸收光谱分别为532~550 nm和457~485 nm[3]。甜菜色素在pH 4~6、4 ℃条件下表现最稳定[4,9]。因此,甜菜色素被广泛应用于低温冷藏食物中,如奶制品[9]。甜菜色素对光温的稳定性不够突出,但是短暂高温有助于提高甜菜色素的稳定性[10]。
甜菜色素稳定性的强弱是由其化学结构决定的。甜菜红素的热稳定性是甜菜黄素的三倍[11]。甜菜色素的稳定性受环境色素浓度、低糖基化、低酰化、PPO和POD等降解酶、金属离子(如Fe3+、Fe2+、Cu2+、Cu+、Sn2+、Al3+、Hg2+、Cr3+等),以及高温、光照、高氧等因素影响而加速降解[12—13]。原理是上述因素诱导胺键断裂、脱氢作用、去糖基化、脱羧作用、异构化等从而导致甜菜色素分子内部结构发生断裂(图2)[14]。为了提高甜菜色素的商业化应用,减少降解最可行的手段是添加一定的抗氧化剂、微囊化、像花青素一样共色素作用等[15—17]。生产上可通过添加抗坏血酸提高甜菜色素的稳定性[16—17]。Otálora等[16]从紫色的仙人掌果实中提取甜菜色素然后通过加入海藻酸钙和牛血清白蛋白进行离子凝胶化稳定,甜菜色素胶囊化技术是提高甜菜色素稳定性的有效方法。
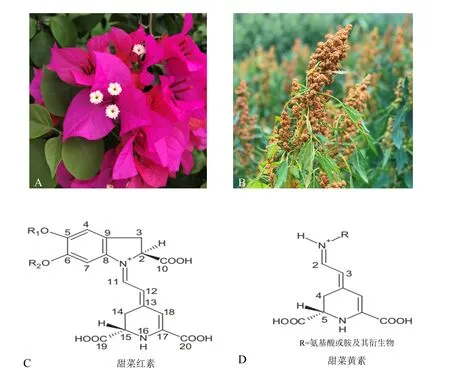
图1 甜菜色素在植物中的色彩及代表性结构Fig.1 Color and representative structures of betalains
2 甜菜色素生理作用及功能性特征
甜菜色素在植物体内不仅使花朵美丽以吸引传粉者繁衍后代,也是实现其花朵和果实经济价值的重要因素[17—18];生理上,它的存在对植物本身抵抗逆境如干旱胁迫和过氧化胁迫也起着重要作用[19—20]。仙人掌科植物刺梨Opuntia ficus生长于干旱地带,它可通过甜菜黄素的合成或降解来改变氨基酸浓度从而进行渗透调节,提高抗旱能力[19]。
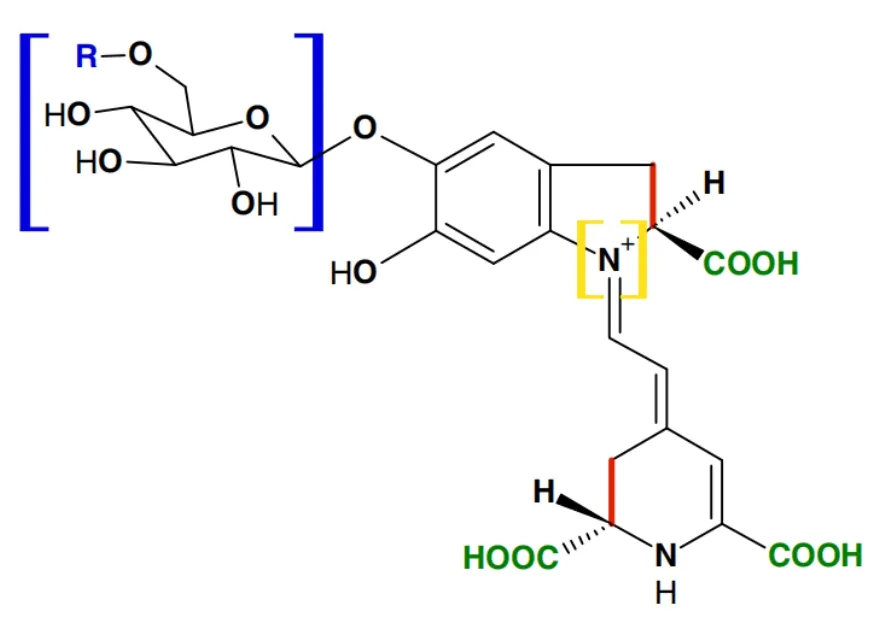
图2 甜菜色素分子的可能降解位置[4]Fig.2 Factors affecting betalain stability[4]
甜菜色素作为重要的植物次生代谢物质,与花色素苷一样也有抗氧化、光保护作用[21],其抗氧化和抗辐射作用比膜保护剂Trolox (维生素E类似物)更显著[22—24]。研究表明,食用仙人掌科刺梨果实一段时间可提高人体抗氧化水平[25];刺梨提取物在体外具有较强的抗菌和抗氧化效果,其机理可能与所含甜菜色素的抗氧化作用有关[19,25]。此外,甜菜色素可通过作用人骨髓内的过氧化物酶(myeloperoxidase)清除次氯酸[26]。由此可见,长期食用含有甜菜色素的食品有助于缓解外部压力引起的身体失调[27—28]。
3 甜菜色素合成
甜菜色素合成代谢的研究始于20世纪90年代,合成途径主要包括酶促反应和自发反应,反应起始于酪氨酸。参与合成途径的酶主要有酪氨酸酶(Tyrosinase)、葡萄糖基转移酶(UDP-glucosyltransferase)、多巴双加氧酶(4,5-DOPA-extradiol-dioxygenase)和细胞色素单氧化酶(Cytochrome P450-like)。甜菜色素合成的第一个底物是L-酪氨酸,酪氨酸酶有双重作用,该酶的单酚氧化酶(Monophenolase)作用产生多巴(L-DOPA)[29—30],另一重活性双酚氧化酶(Diphenolase)导致多巴氧化成多巴醌(o-DOPA-quinone)[31],这样消耗新生成的L-DOPA,而细胞中的抗坏血酸或者类似物可协助多巴醌还原成多巴[14,17,31]。酪氨酸酶普遍存在于动植物中,在动物体内该酶促成色素的形成;在植物体内,除了甜菜色素代谢类型植物中是甜菜色素合成途径的重要酶外,在花青苷代谢途径的植物中也有发现,但功能还不很清楚[32]。酪氨酸酶具有单酚和双酚氧化酶的双重功能,属于PPO类型酶。该酶最早从毒蝇伞中纯化的[33],之后又从大花马齿苋Portulaca grandiflora、甜菜Beta vulgaris和盐地碱蓬Suaeda salsa中分离获得[14,18,29—30,34]。
甜菜色素合成途径的第二个关键酶是多巴双加氧酶,该酶催化 L-DOPA形成中间产物开环多巴(4,5-seco-DOPA)。该酶首先是从毒蝇伞中鉴定分离的[35—36],之后也从大花马齿苋和甜菜中分离出来[37]。形成开环多巴之后进行自发反应形成甜菜色素的生色团甜菜醛氨酸(Betalamic acid)[14],之后该生色团和各种氨基酸发生自发的缩合反应形成各类甜菜黄素[38]。酪氨酸酶和多巴双加氧酶两个关键酶都属于氧化酶,催化过程依赖于氧分子。
甜菜色素合成途径的第三个关键酶是细胞色素单氧化酶(Cyt P450),该酶催化多巴形成环状多巴(Cyclo-DOPA)[39],其作用机制尚不清楚,推测与电子传递有关。
甜菜色素合成过程中的第四个关键酶葡萄糖基转移酶(UDP-G)则是修饰酶,把糖基转移至甜菜苷配基上,形成甜菜苷元[40]。环状多巴在葡萄糖基转移酶作用下形成环多巴葡萄糖,继而和甜菜醛氨酸自发反应形成甜菜苷(Betanin),这是甜菜红素形成的基本苷元。另一种可能是环状多巴首先和生色团甜菜醛氨酸发生自发反应形成甜菜苷配基,然后在葡萄糖基转移酶作用下形成甜菜苷[41]。从番杏科彩虹菊Dorotheanthus bellidiformis中分离纯化出葡萄糖基转移酶[42—43]。
甜菜红素和甜菜黄素的合成过程受几个关键酶的调控程度有较大差异。甜菜黄素的积累取决于一个分支途径,而甜菜红素的合成由多个途径完成。甜菜黄素的合成直接正相关于酪氨酸酶和多巴双加氧酶的表达强度;甜菜红素的形成由于受多个分支的影响,细胞色素单氧化酶和葡萄糖基转移酶表达强度与甜菜红素积累的线性关系不显著[44]。
目前,大自然中未发现两类色素共存于同一种植物,但是越来越多的表观遗传学和反向遗传学研究发现,花青苷代谢途径的酶和转录因子可以在甜菜色素代谢类型的植物中表达;反之,甜菜色素代谢途径的酶和转录因子也可以在花青苷代谢植物中表达并翻译成相应的蛋白[44—50]。
4 甜菜色素的应用
由于甜菜色素的保健价值日益被人们所熟知,加上其较高的酸碱稳定性,近年来被越来越看好开发成天然食用色素用于食品工业[10]。甜菜色素的开发符合食品添加剂国际需求潮流:安全、保健的功能型食品添加剂。2009年统计数据表明,全球每年对食用色素的需求量达到45 000~50 000 t[12],其中对红甜菜来源的甜菜色素的需求占10%以上[51]。目前,欧盟已经批准甜菜红素用于食品(EEC No.E162)[52],美国药品食品管理局(FDA)也正式批准甜菜红素在食品领域的使用(No.73.40)[53],这类色素由于在高pH(>4)环境中更稳定而可能被更广泛应用(其抗氧化能力为花青苷色素的1.5~2倍)。甜菜色素的适用范围包括糖果、酸奶、沙拉、软饮料等[5],添加50 mg·kg-1甜菜色素即可有良好的着色效果。甜菜黄素由于含有特殊氨基酸,被赋予独特于甜菜红素的理化性质和保健价值,将成为食品中补充氨基酸的重要来源。目前市场上还没有商品化的甜菜黄素销售,原因一是来源较少,二是其弱稳定性。红甜菜是商业化甜菜色素的主要来源,但来自于红甜菜的甜菜红素由于来源部位位于土壤中容易携带微生物,而且带有强烈的土腥味。市场上对于其他甜菜红素来源代替红甜菜有强烈的需求。目前,火龙果Hylocereus undatus由于不具有上述红甜菜的劣势有望成为甜菜红素的新来源,但是其果胶类物质过多。因此,开发更多优良的甜菜色素来源是扩大其应用的根本措施。
