毛地黄鼠尾草花部性状的变异及其对传粉昆虫访花偏好的影响
2019-03-22孙淑范骆望龙张勃
孙淑范,骆望龙,张勃
(甘肃农业大学草业学院,甘肃 兰州 730070)
1859年,达尔文[1]提出植物花丰富的多样性是自然选择不断积累的结果[2].许多研究表明,被子植物的繁殖特征,尤其是虫媒传粉植物花部表型特征丰富的多样性在很大程度上是由传粉者介导(pollinator-mediated)的自然选择而引起[3-5].因此,研究植物种群内花性状的变异性及其与传粉者的互作关系成为理解植物花的多样性分化及其进化意义的重要途径.植物受传粉者选择驱动的花表型进化中,有2类性状比较关键:一类是与植物的花吸引力有关的性状,如花大小、花展示和花报酬等[6],这些性状能通过影响传粉者访花频率影响个体的适合度,从而容易受到选择;另一类是花部结构性状,特别是雌、雄蕊的空间布置(如柱头位置)以及花冠结构等,这些性状能通过与传粉者的机械互作影响传粉效率而受到选择[7].因此,这2类性状通常作为植物花表型适应性及其功能研究的目标性状.
鼠尾草(Salvia)是唇形科(Labiatae)最大的一个属,有近1 000个物种,广泛分布在新旧世界[8-9].绝大多数鼠尾草属物种具有高度特化的杠杆状雄蕊结构(lever-like stamens)[10],因其与传粉昆虫互作形成了精巧的背部传粉机制(dorsal pollination),被认为是激发该属植物物种分化和适应性辐射过程的
关键性状(key innovation)[11].因此,鼠尾草属植物成为了花适应性进化研究的理想类群之一[12].本研究以分布在我国青藏高原横断山区的毛地黄鼠尾草(S.digitaloides)为对象,通过观测花性状的变异性及其对传粉昆虫访花行为的影响,探讨了该属植物花表型的适应性及可能的进化生态学过程,以期回答如下2个问题:在自然居群中,毛地黄鼠尾草的花表型性状的变异程度有多大,主要发生在个体间还是个体内;花表型的变异对其传粉者的访花行为是否产生影响,即传粉者是否存在访花偏好.
1 材料与方法
1.1 花部性状测量与结果
本研究以分布在滇西北丽江玉龙雪山的毛地黄鼠尾草为试材.花部性状的测量,参照张勃等[13]的方法.盛花期,随机选择毛地黄鼠尾草个体(>150),用电子游标卡尺测量其花性状.测量性状包括雄蕊杠杆长度、下臂高度、花冠长、冠腹高、冠口高、冠口宽以及柱头高度、柱头水平距离和柱头探出花冠距离9个花部结构性状,如图1所示.同时,统计单株小花数并测量其花序长.每株测量4~5朵花,其平均值作为单株花性状的表型值.

A:花的侧面图;B:雄蕊杠杆放大图;C:花冠口正面图;Ceh: 冠口高;cew:冠口宽;cl:花冠长;ct:冠腹高;lah:杠杆高;se:柱头探出距离(柱探);shd:柱头水平距离(柱平);sll:雄蕊杠杆度;sth:柱头到花冠口下沿的距离(柱高).A:Side view of a flower;B:Enlarged view of stamen lever;C:Front view of a flower;Ceh:Corolla entrance height;Cew:Corolla entrance width;cl:Corolla length;ct:Corolla thickness;lah:Stamen lower arm height;se:Style exsertion;shd:Stigma horizontal distance from lower arm;sll:Stamen lever length;sth:Stigma height.图1 毛地黄鼠尾草的花形态及其花性状测量Figure 1 Flower construction andmorphometrics in Salvia digitaloides
1.2 传粉者及其访花偏好观测
在毛地黄鼠尾草种群的盛花期,调查所有的访花昆虫,并带回实验室鉴定.访花观察期间,随机跟踪正在访花的传粉昆虫,观察其访花行为.如果传粉昆虫访问一朵花后,紧接着飞向另一朵花,但是当其接近下一朵花的花冠口后,随即又飞离,被视为访花有偏好.若这两朵花属于同一株,被认为是株内访花偏好;若属不同株,则被认为是株间访花偏好.在此期间,同时摘取拜访花和下一朵未访花,测量其花部性状,包括花冠长、花冠口大小以及雄蕊的下臂高度4个性状.
1.3 花蜜产量测量
在盛花期,随机选取含苞待放的花(>100)进行套袋.套袋次日,选择已开放的花,待24 h后,同时测量其花蜜产量和花部结构性状(包括花冠长、冠口宽、冠口高和雄蕊下臂高).花蜜量用毛细管法进行测量.
1.4 数据统计
所有数据用R软件[14]进行统计分析.花蜜产量与花部性状的关系以及传粉者拜访花与未访花之间的差异性(即访花偏好)用R 软件一般线性模型(LM)完成.
2 结果与分析
2.1 毛地黄鼠尾草花部性状及其变异性
毛地黄鼠尾草的花性状表型值及其变异系数如表1所示.总体上,该物种花性状在不同个体间的变异程度明显大于个体内,各性状之间的变异性存在明显差异,在所测量的12个花性状中,花蜜量的变异系数最大,为44.99,小花数次之,为29.46.在花部结构性状中,有关柱头位置性状的变异程度相对较大,柱高、柱头水平距离与柱头探出距离在个体间的变异系数分别为18.90、19.07和24.34;其他花部结构性状如冠长、冠口大小的变异程度最小其变异系数小于11%.
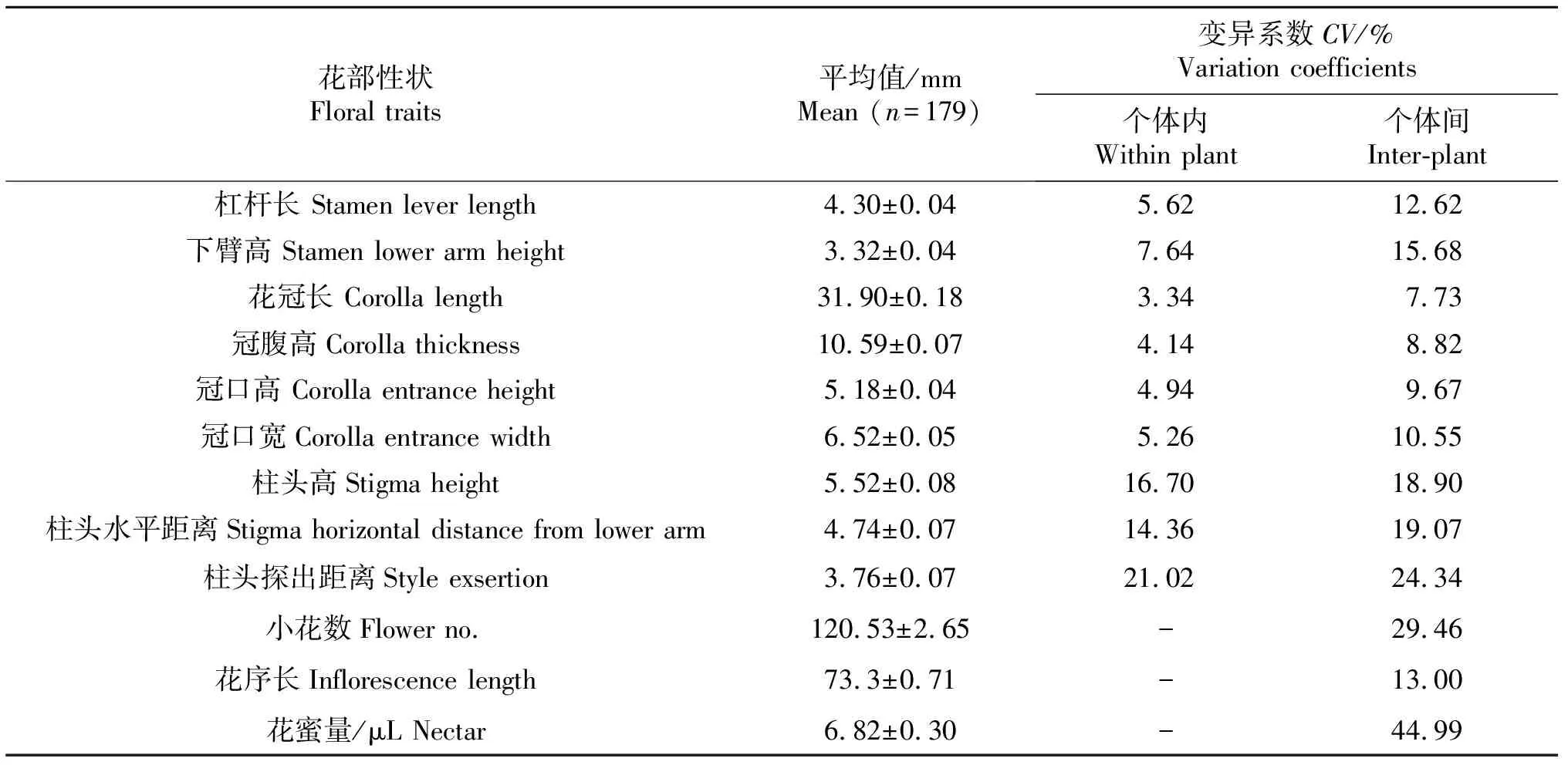
表1 毛地黄鼠尾草的花性状及其变异系数
2.2 传粉昆虫及其访花偏好
毛地黄鼠尾草种群的传粉者包括弗里熊蜂(Bombusfriseanus)、稀熊蜂(B.infrequens)和伪猛熊蜂(B.personatus)3个物种.弗里熊蜂占所有熊蜂访花频率的65%,是其主要的传粉者.该熊蜂在访花时,对不同性状的花存在明显的访花偏好,结果如图2所示.当传粉者在同株(花序)内访花时,拜访花的平均花冠长、冠口宽和冠口高分别为(32.58±0.29) mm、(6.73± 0.08) mm和(5.25± 0.06) mm,显著大于同期未访花的性状值(图2-A~C,n=116,P<0.001),而同期拜访花与未访花的下臂高无显著差异(图2-D,n=116,P=0.117).当传粉者在不同个体间访花时,其拜访花与未访花的花冠长、冠口宽、冠口高以及下臂高无显著差异.

图2 毛地黄鼠尾草传粉者拜访花与未访花的花部性状比较Figure 2 Difference in floral traits between probed and nou-probed flowers in S.digitaloides
2.3 花蜜量与花部结构性状的关系
毛地黄鼠尾草花部结构性状与花蜜产量之间的关系如图3所示.在所测量的花性状中,花冠长(图3-A)、冠口宽(图3-B)、冠口高(图3-C)及冠腹高(图3-D)与花蜜产量之间均具有明显相关性.其中,花冠长和冠腹高与花蜜产量具有极显著的正相关关系(n=106,P=0.005,0.002),相关系数分别为0.274和0.295.冠口大小,即冠口宽和冠口高与花蜜产量存在近显著的相关关系,二者的相关系数分别为0.187和0.197.其余花部性状,如杠杆长、下臂高、柱高、柱平和柱探等与花蜜产量之间不存在相关性.
3 讨论
花是被子植物的繁殖器官,其丰富的多样性被认为是通过自然选择不断积累的结果[1].由达尔文自然选择理论模型可知,植物与其适合度相关的性状存在可遗传变异是该物种通过自然选择不断进化的基础.研究发现,植物花39%的变异为可遗传变异,且不同花性状具有不同的遗传力[15].本研究表明,毛地黄属尾草的花部性状无论在个体内还是个体间均存在很大的变异性,各性状的变异程度也表现不同.所测量的花性状中,花蜜量的变异程度最大,花数量次之;花部结构性状中,有关柱头位置的花性状,如柱头探出距离和柱头高度,其变异性明显高于其它花结构性状.这与Gresswell[16]的研究结果相类似.总体上,毛地黄鼠尾草的大部分花性状在个体内的变异程度明显小于个体间,表明该物种的花表型性状尽管存在较大的可塑性,但个体间相对更大的遗传变异反映出该物种能通过自然选择而不断进化的潜力.
植物的花与其传粉者的互作是被子植物进化的重要推动力[17-18].一方面,花部特征(如花报酬)对传粉者具有吸引作用;另一方面,花部结构对传粉者有机械适应性.因此,花部性状的变异能引起传粉效率的个体差异而受到选择,即花表型会在传粉者驱动下向适应其传粉实现的特定方向进化[19].研究发现,很多植物的花部性状,如花大小和花蜜量等,能引起传粉者的访花偏好而受到强烈的选择[20-21].本研究表明,毛地黄鼠尾草的花蜜产量与花冠长、花冠口大小和花冠腹高度3个性状之间存在显著的正向相关性;相应地,传粉昆虫(弗里熊蜂Bumblefriseanus)对较长花冠和较大冠口的花表现出明显的访花偏好.一方面,传粉昆虫可能会通过花(冠口)大小区分花报酬的多少,故而对不同大小的花表现出访花偏好;另一方面,鼠尾草特化的雄蕊杠杆花结构对传粉昆虫有一定的阻拦和筛选作用[12,22],相对较小的花,尤其是小冠口花能增加传粉者的访花代价,相反的,大冠口的花更易于吸取花蜜而受到传粉者的青睐,因此,传粉者对相对较大的花表现出访花偏好.另外,本研究显示,传粉熊蜂的访花偏好主要表现在植株个体内不同的花之间,而并非不同个体间,这很可能是由于传粉熊蜂访花记忆能力的局限性所致[23].也就是说,访花期间,当熊蜂在鼠尾草不同个体间移动时,由于耗费时间相对较长,会丢失对不同花大小的比较记忆,从而在不同个体间未表现出明显的访花偏好.
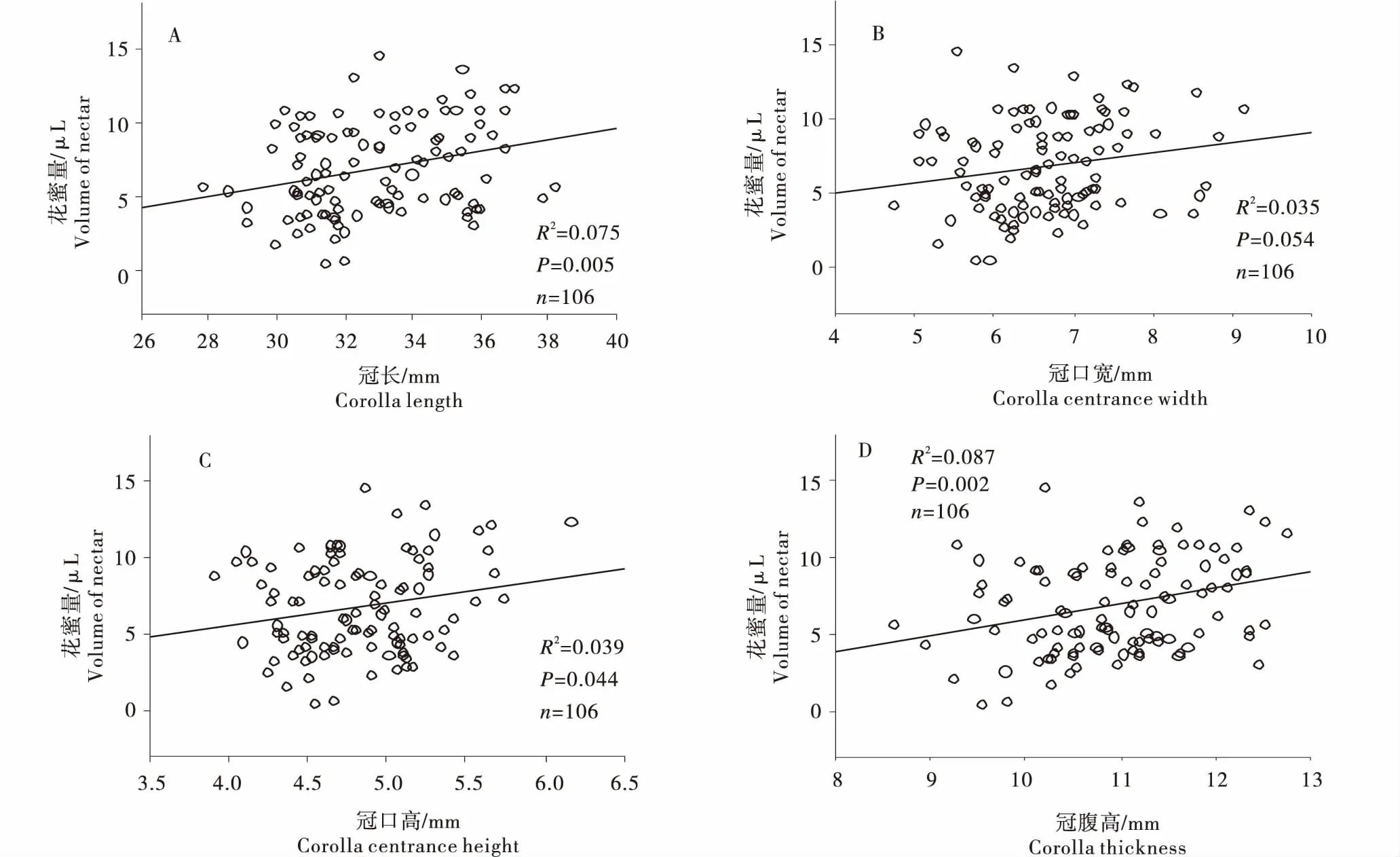
图3 不同花性状与花蜜量的关系Figure 3 Relationship ofnectar productionwith floralstructural traits in S.digitaloides
