青藏高原不同海拔高度下全缘叶绿绒蒿叶表皮特征研究
2019-03-19李苗苗唐道城
唐 楠 李苗苗 唐道城*
(1.青海大学高原花卉研究中心,西宁 810016; 2.青海省园林植物与观赏园艺重点实验室,西宁 810016)
全缘叶绿绒蒿(Meconopsisintegrifolia(Maxim.) Franch.)是罂粟科(Papaveraceae)绿绒蒿属(Meconopsis)一年生至多年生草本植物,主要分布于我国西藏、青海、四川、甘肃及云南西北部,通常生长于海拔3 000 m以上的流石滩、高山灌丛和雪山草甸,具有较强的抗旱、抗寒及耐瘠薄能力,在药用和观赏方面具有较高的开发利用价值[1~2]。目前,对于全缘叶绿绒蒿的研究较少,主要集中在植物化学成分和药理方面[2~5],对全缘叶绿绒蒿的传粉生态学、适应进化等方面的了解较少。吴云等[6]在不同海拔高度下对全缘叶绿绒蒿的传粉生态学进行了观察,探讨了全缘叶绿绒蒿的繁育系统特点和其对高山气候环境的繁殖适应特征。张霓雯等[7~8]对不同海拔下全缘叶绿绒蒿的繁殖分配特征、种子散播功能性状、种群空间分布格局特征进行了研究,探讨了全缘叶绿绒蒿繁殖分配策略对海拔的响应,以及种子散播功能性状与种群空间分布格局特征的相关性。在叶片解剖结构方面,刘梦颖等[9]对四川省汶川县卧龙保护区巴郎山的全缘叶绿绒蒿的叶片形态及解剖结构进行了研究,为了解全缘叶绿绒蒿对海拔及高山环境的形态学适应特征奠定了基础。
植物进化过程中,叶片结构对环境变化较为敏感,其结构特征最能体现环境因子的影响以及植物对环境的适应性[10]。青藏高原是中国最大、世界海拔最高的高原,具有独特的气候特征,如辐射强烈、日照多、气温低、积温少、氧气稀薄等[11]。生长在高原环境的植物叶片结构常表现出一些旱生植物的特征,如气孔多分布在叶片下表皮,形成气孔窝,气孔面积趋小,密度趋多[12~13],栅栏组织多层且发达,海绵组织减少或消失等特征[14]。表皮毛是由植物表皮细胞发育而来,生长在表皮组织的一种特化结构[15~16]。表皮毛与表皮上的气孔、角质层、蜡质等表皮特征互相配合,共同完成各种不同的保护功能[17~18]。植物为了适应生长环境,表皮毛会对盐分、干旱、海拔和光照等环境因子作出响应[19~21]。有关海拔高度对植物叶片解剖结构影响的研究较少,施海燕等[22]对青藏高原东缘海拔2 100~4 200 m处生长的7种风毛菊属植物的叶结构进行了显微观察,分析探讨7种植物叶解剖结构与自然环境的相互关系。孙会婷等[23]对分布于青藏高原祁连山东部不同海拔高度的蒲公英、火绒草和美丽风毛菊的叶片形态解剖结构进行了研究,认为3种植物采取不同的响应机制来适应海拔高度的变化,植物对高原环境变化的适应具有多样性。本文以分布于青藏高原5个不同海拔高度下的全缘叶绿绒蒿基生叶为研究材料,观测叶片的表皮毛、气孔及表皮细胞结构特征,分析叶片表皮毛、气孔器和表皮细胞特征与生态环境的关系,旨在探讨全缘叶绿绒蒿叶片表观特征及解剖学对高原生态环境的适应性响应。
1 材料与方法
1.1 实验材料
实验材料为2007年7~8月采自青藏高原东北部的高山地区(海拔3 500~4 800 m,97°39′~105°26′E,33°54′~35°13′N)的全缘叶绿绒蒿。采样点的生境信息详见表1。每个样点随机选取5~10株全缘叶绿绒蒿,每株相同部位摘取健康的基生叶5片,部分装入塑料自封袋中低温保存鲜样,其余剪取叶的中央部分1.5 cm×1.5 cm,用50%酒精配制的FAA固定液固定。

表1 采样点生境信息
1.2 实验方法
1.2.1 上下表皮毛密度观测
每样点取开花植株5株,每株取5片基生叶,每片基生叶在XTT型实体解剖镜下直接观察并记录表皮毛数量,上下表皮各观测5个视野。用单位面积的表皮毛数量表示表皮毛密度,即:
表皮毛密度(个·mm-2)=视野平均表皮毛数/视野面积
(1)
1.2.2上下表皮气孔器和表皮细胞密度及形态观测
采用胶带粘取法[24]制作上下表皮临时装片。在Nikon光学显微镜10×10倍下观察气孔器形态、记录气孔器数量,用单位面积的气孔数量表示气孔密度。有关气孔类型的术语参照Dilcher[25]。采用鼠标点测法[24]和相应的计算公式获得气孔长度、宽度以及气孔面积(气孔长度是气孔关闭状态下肾形长度;气孔宽度是垂直于气孔器长轴的最宽值)。取基生叶5枚,每枚叶片制作5张切片,每张切片随机选取5个清晰视野,分别测量相关指标,取平均值作为该叶片的相关数据。表皮细胞测定方法与气孔器相同。气孔密度和表皮细胞密度的计算公式如下:
气孔密度(个·mm-2)=视野平均气孔数/视野面积
(2)
表皮细胞密度(个·mm-2)=视野平均细胞数/视野面积
(3)
2 结果与分析
2.1 全缘叶绿绒蒿叶片表皮毛与海拔高度的关系
观察结果表明,基生叶上表皮的表皮毛分布比较均匀,下表皮的表皮毛在叶片叶脉处分布密度较大。随着海拔高度增加,上、下表皮的表皮毛密度显著增大,而且上表皮的表皮毛分布密度明显大于下表皮(表2)。
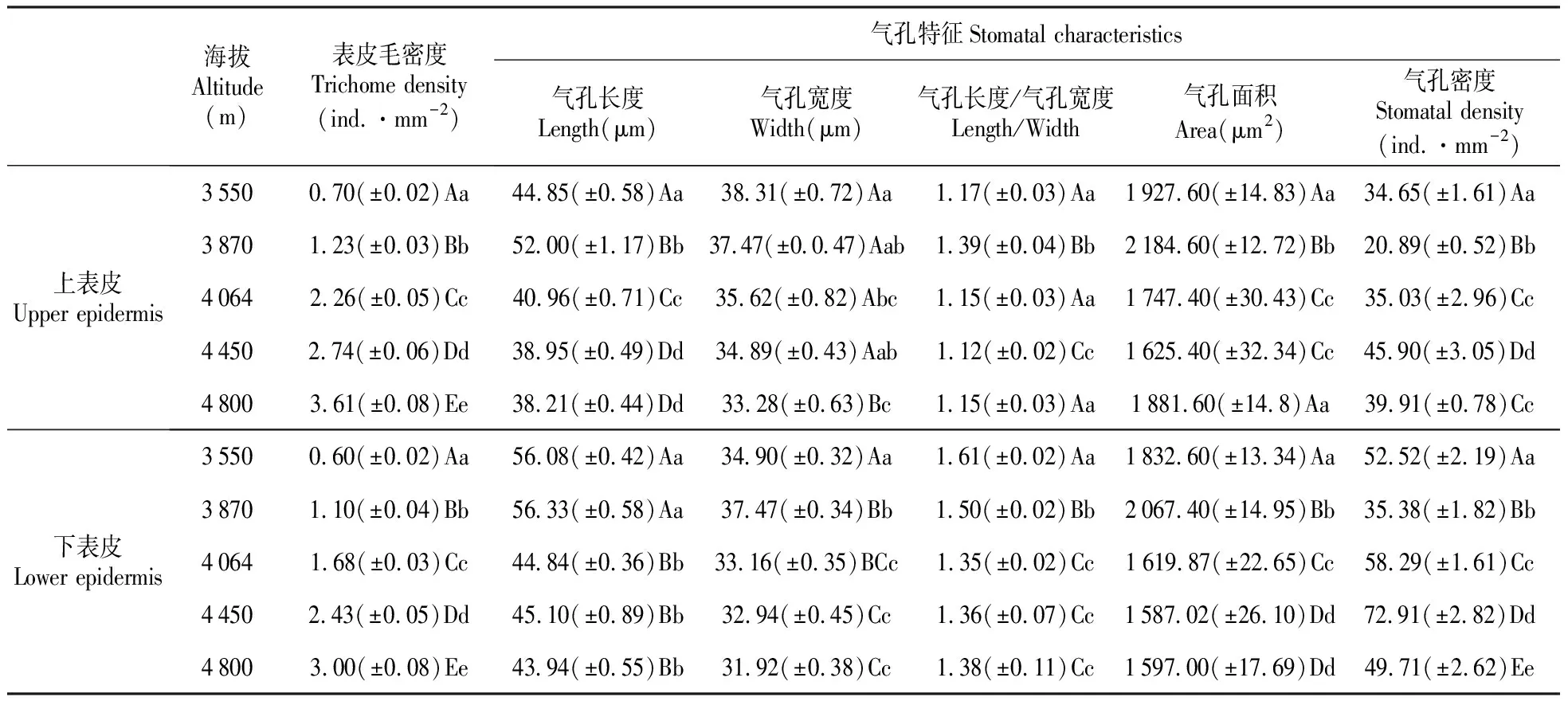
表2 不同海拔高度下全缘叶绿绒蒿叶片表皮毛和气孔变化特征
注:大写字母表示0.01水平的显著性,小写字母表示0.05水平的显著性。下同。
Note:Capital letters represent significance on 0.01 level,and lowercase letters represent on significance on 0.05 level.The same as below.
2.2 全缘叶绿绒蒿叶片气孔特征与海拔高度的关系
全缘叶绿绒蒿的基生叶片上、下表皮的气孔仅由保卫细胞组成,无副卫细胞,由数目可变的普通表皮细胞不规则的围绕在气孔周围(图1)。气孔类型均为无规则型;上表皮气孔器近于椭圆形,下表皮气孔器近于纺锤形。由表2可知,全缘叶绿绒蒿上、下表皮的气孔长度随海拔升高而逐渐变短,上表皮气孔长度<下表皮气孔长度;随着海拔的升高,气孔宽度逐渐减小,上表皮气孔宽度>下表皮气孔宽度。在海拔3 870 m,上、下表皮的气孔长度达到最大;在海拔3 870~4 450 m,上、下表皮的气孔长度急剧缩短,当海拔高于4 450 m时,上、下表皮的气孔长度稳定保持在较短水平,上表皮的气孔长度/气孔宽度均小于下表皮。上、下表皮的单个气孔面积随海拔的增高逐渐减小,上表皮的单个气孔面积>下表皮的单个气孔面积,其变化规律与气孔长度变化规律基本一致。上、下表皮的气孔密度变化趋势与气孔面积变化趋势相反,即随着海拔高度的增加,气孔密度逐渐增加,上表皮的气孔密度<下表皮的气孔密度。
2.3 全缘叶绿绒蒿叶片的表皮细胞特征与海拔高度的关系
随着海拔高度增加,叶片的上、下表皮细胞形状、大小及垂周壁式样均发生明显变化(图1,表3)。在海拔4000 m以下,叶片的上、下表皮细胞呈无规则形,垂周壁呈波状;在海拔4 000 m以上,叶片的上、下表皮细胞呈多边形,垂周壁呈弓形或平直;并且随着海拔高度增加,叶片的上、下表皮细胞逐渐演变至近圆形,垂周壁逐渐演变成圆弧形。研究结果还显示,叶片表皮细胞的各特征参数也随海拔高度增加也呈现一定的规律性变化,即上表皮细胞长度<下表皮细胞长度,上表皮细胞宽度﹤下表皮细胞宽度,上表皮细胞面积>下表皮细胞面积,上表皮细胞密度<下表皮细胞密度,上表皮细胞长度/细胞宽度<下表皮细胞长度/细胞宽度(表3)。此外,随海拔高度增加,上、下表皮细胞长度、宽度和面积的变化趋势与气孔长度、宽度和面积的变化情况相一致。

表3 不同海拔高度下全缘叶绿绒蒿叶表皮细胞变化特征
3 讨论
青藏高原是绿绒蒿属植物的集中分布区,其总量占绿绒蒿属植物的15%左右[1],全缘叶绿绒蒿是绿绒蒿属的一个种。青藏高原生态环境复杂,气候多变,空气稀薄、O2和CO2分压低,太阳辐射量大、短波辐射(紫外线)强,劲风频发,昼夜温差大、气候寒冷,降水量少,大气及土壤干旱严重[26],而这些环境因子很大程度随海拔高度的变化而随之发生变化。海拔剧变对这些环境因子的综合作用必然会对植物的分布、生长及发育产生一定的影响。叶片是植物进化过程中对环境变化较敏感,而且是可塑性较大的器官,对环境的反应通常较多的反映在叶片的表观特征及解剖结构上,其中解剖结构及微观特征最能体现环境因子的影响或植物对环境因子的响应[27~29]。
本研究结果表明,随着海拔高度的增加,全缘叶绿绒蒿叶片上、下表皮毛的密度逐渐增加,这与段喜华等[30]对不同海拔泡沙参(Adenophorapotaninii)的研究结果相似。表皮毛增加了植物表皮厚度,为植物提供了一道天然的物理屏障,减轻生物和非生物胁迫对植物的伤害[31]。表皮毛的密度增大,减少了高海拔强气流对气孔的影响,降低蒸腾,同时有效防止强光直射对植物的伤害。随着海拔增加,叶片的上、下表皮毛密度逐渐增加,这是植物对高山环境中的强辐射以及寒冷气候的适应性表现。
本研究中,总体上叶片上、下表皮气孔大小参数随海拔升高而逐渐减小,气孔密度逐渐增加,但有在海拔3 870 m处出现折点的现象。一般情况下,随海拔上升CO2分压降低,在低CO2分压下,气孔密度会升高,增加气孔吸收CO2面积,从而保证光合作用的进行[32]。在3 870 m出现折点,可能是因为在增强气孔吸收面积的同时,增强了水分蒸发,这在高海拔的低温、大风环境中更易造成植物的“生理干旱”[33],因而气孔特征参数出现折点,这与李芳兰[27]、贺金生[28]、马书荣等[34]的研究结论一致。然而,也有研究表明蒲公英和美丽风毛菊的上、下表皮气孔密度则随海拔的增加而降低[23],这可能是不同植物对环境因子的敏感性不同所致,这两种植物对环境变化十分敏感,海拔高度增加,气温下降,植物通过降低气孔密度来适应低温环境[35]。
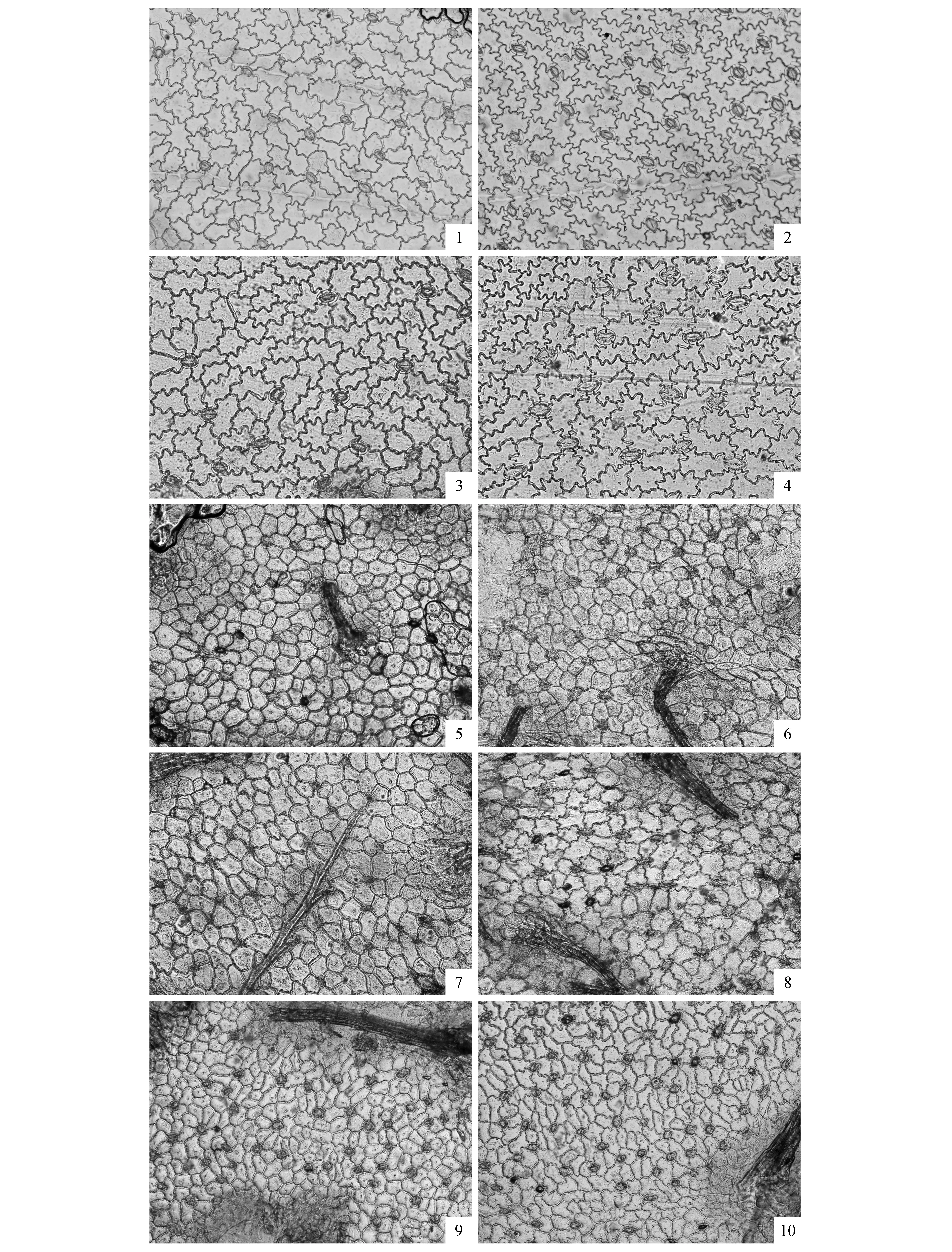
图1 光镜下全缘叶绿绒蒿叶片表皮显微结构 图1、3、5、7、9分别表示海拔3 550、3 870、4 064、4 450、4 800 m时的全缘叶绿绒蒿上表皮显微结构;图2、4、6、8、10分别表示海拔3 550、3 870、4 064、4 450、4 800 m时的全缘叶绿绒蒿下表皮显微结构。Fig.1 Morphology of M.integrifolia leaf epidermis structure under LM Figure 1,3,5,7,9 represent upper leaf epidermis structure under altitude 3 550,3 870,4 064,4 450 and 4 800 m; Figure 2,4,6,8,10 represent lower leaf epidermis structure under altitude 3 550,3 870,4 064,4 450 and 4 800 m
本文对全缘叶绿绒蒿叶片上、下表皮细胞的大小及密度变化进行了观测和分析,其变化趋势与气孔特征参数的变化趋势基本一致。前人曾对不同海拔下全缘叶绿绒蒿的叶片表皮厚度进行了观测,结果表明叶表皮细胞厚度随海拔的升高而减小[9],但没有对叶片表皮细胞面积和密度进行观测。本研究发现叶片上、下表皮细胞的大小总体上随海拔增高而减小,这可能是受高海拔环境胁迫,表皮细胞控水能力受阻,根系吸收的水分不能满足蒸腾散失的水分,导致植物缺水,出现“生理饥渴”,为了适应这种情况,叶片会发生表皮细胞变小,细胞壁加厚,角质膜增厚等变化[36]。尽管环境因子变化对叶表皮细胞的影响研究报道较少,但通过对不同海拔全缘叶绿绒蒿表皮细胞特征变化规律的研究,可对于以后的研究提供借鉴。
本文通过对青藏高原5个不同海拔高度下全缘叶绿绒蒿叶片表皮毛、气孔和表皮细胞特征变化规律的研究,对于揭示全缘叶绿绒蒿与生态适应性之间的关系和对环境变化的响应及其适应机理奠定了一定的理论基础,有助于对揭示该物种的生态演变及在未来气候环境变化条件下的保护和开发应用都具有重要意义。
