疾跑时股后肌群的生物力学特征
——对拉伤危险时相的分析
2019-03-06万祥林
万祥林,曲 峰,刘 卉,于 冰
股后肌群拉伤是体育运动中最为常见的损伤之一,多发于田径、足球、澳式橄榄球、英式橄榄球、美式橄榄球等涉及快速奔跑、跳跃、踢等动作的项目(Mendiguchia et al.,2012;Opar et al.,2012),其中,疾跑是最主要的损伤动作形式(Brooks et al.,2006;Woods et al.,2004)。拉伤的高损伤率和复发率会导致运动员缺席比赛,造成巨大的经济损失等(Askling et al.,2006;Ekstrand et al.,2012;Hickey et al.,2014)。尽管大量的人力、物力、财力被投入用于预防股后肌群的拉伤和提高拉伤的康复效果,然而近30年来,股后肌群的损伤率一直没有下降(Liu et al.,2012;Ramos et al.,2017),因此,当前迫切需要研究并提出预防拉伤和提高康复效果的有效措施。
明确损伤机制是预防股后肌群拉伤和提高损伤后康复效果的前提,而了解疾跑时股后肌群拉伤的生物力学条件对于探明拉伤机制具有重要意义。目前,针对股后肌群拉伤易发生在跑步周期的支撑相还是摆动相仍存在争议。由于人的痛觉的滞后性,很难通过现场视频或伤者本人的描述来判断运动中拉伤发生的时刻,因此,实验室生物力学研究是分析跑动过程中股后肌群易伤时相的主要方法。Mann(1981)和魏书涛等(2009)发现,屈膝力矩和伸髋力矩的峰值出现在疾跑支撑相初期,因此认为股后肌群承受巨大负荷使得支撑相初期成为拉伤发生的高危时相。Sun 等(2015)还发现,在摆动相末期,有一个比支撑相初期更大的屈膝、伸髋力矩,认为支撑相初期和摆动相末期均是疾跑股后肌群拉伤的高危时刻。然而,许多学者通过动物模型研究表明,拉伤主要是在肌肉处于拉长状态下发生的,拉伤的直接原因是肌肉应变过大,而与肌肉收缩力和速度不相关(Yu et al.,2017),因此,屈膝、伸髋力矩大并不一定是股后肌群拉伤的原因。肌肉应变是指肌肉相对静息长度的增量与肌肉静息长度的比值(Enoka,2008)。Thelen 等(2005)发现,在跑台上疾跑的摆动相末期,股后肌群出现长度峰值且处于离心收缩状态,故认为拉伤易发生在摆动相末期。Yu等(2008)分析了地面上跑时股后肌群的运动学及肌电特征,发现在摆动相末期和支撑相末期,股后肌群均出现长度峰值并处于离心收缩状态,且摆动相末期的长度更长、激活程度最高,因此得出疾跑的摆动相末期和支撑相末期是股后肌群易拉伤的时相,且摆动末期拉伤的风险可能更高。这些分析运动中股后肌群拉伤机制的研究,或是采用股后肌群的绝对长度,或是采用肌肉长度相对直立时肌肉长度的形变量来间接反应肌肉的应变,鲜见讨论与拉伤直接相关的股后肌群应变特征。
本研究在测量计算受试者股后肌群最优长度的基础上,分析疾跑一个步态周期过程中股后肌群应变特征,同时结合疾跑过程中下肢运动学、动力学和股后肌群激活特征等参数的分析,探讨股后肌群拉伤的危险时相,深入分析疾跑时股后肌群拉伤的机制。
1 研究对象与方法
1.1 受试者
本研究受试者为男、女各20名有短跑训练经验的大学生(表1)。所有受试者无股后肌群拉伤史,下肢及足部半年内无明显损伤,实验前48 h未从事剧烈运动。所有受试者均签署了知情同意书。
1.2 数据采集与处理
受试者着紧身衣裤,在身上粘贴反光标志点(第4、5腰椎棘突中点,以及左右侧的髂前上棘、髂嵴最高点、股骨大转子、股骨外侧髁、股骨内侧髁、胫骨粗隆、腓骨外踝、胫骨内踝、足跟及足尖),通过肌电仪(NoraxonTeleMyo 2400T G2,Noraxon Inc.,Scottsdale,AZ,USA,3 000Hz)采集两侧股二头肌和半腱半膜肌的肌电信号,同时通过8镜头红外高速运动捕捉系统(Motion Analysis Raptor-4,Motion Analysis Corp.,Santa Rosa,CA,USA, 200Hz)、三维测力台(Kistler 9281CA,Kistler Instruments,Winterthur,Switzerland,1 000 Hz)与肌电同步采集体表标志点坐标和地面反作用力数据。跑步距离约35 m,受试者在距离测力台直线距离约20 m处以站立式起跑姿势准备,听到口令后尽最快的速度跑过测试区域。受试者以正常步态、无任何步伐调整并以测试脚着在测力台上视为1次有效测试。受试者每侧腿完成3次有效测试,每次之间休息60 s,疾跑测试获得的指标取3次结果的平均值。

表1 受试者基本信息Table 1 Basic Information of Subjects
受试者完成疾跑测试后,保持原有体表标志点不变,坐在等速测试仪上(Isomed 2000,D&R Ferstl GmbH,Hemau,Germany),保持髋屈约90°,测试腿的大腿及小腿分别被固定在座椅和机械臂上,防止代偿动作以保证只能完成膝的屈伸运动,膝的屈伸轴与机械臂的转动轴对齐。测试的机械臂活动范围为110°,膝完全伸直时机械臂的位置为0°。受试者每侧腿尽全力完成3次角速度为10°/s的等速屈膝运动,每次之间休息90 s。体表标志点的坐标通过10镜头红外运动捕捉系统(Qualisys,Gothenburg,Sweden)采集,并与等速力矩数据同步。
采集的所有体表标志点三维坐标,运用Butterworth低通滤波法进行平滑,截断频率为13 Hz(Yu et al.,1999)。根据标志点坐标建立骨盆、大腿、小腿和足的环节坐标系。参照前人研究(Wan et al.,2017;Yu et al.,2008)计算股后肌群各肌肉起、止点在大地坐标系中的坐标。肌肉的长度为大地坐标系中肌肉起、止点连线的距离。由于目前难以获取在体肌肉的静息长度,肌肉的最优长度可近似当作肌肉的静息长度(Kaufman et al.,1991),肌肉的最优长度是指肌肉产生最大收缩力时的长度(Chang et al.,1999)。股后肌群每块肌肉的收缩力根据等速屈膝力矩、肌肉的生理横截面积、肌肉力臂计算获得,具体的计算方法可参照我们之前的研究(Wan et al.,2017)。某一时刻某块肌肉的应变(εt)由这一时刻肌肉的长度(Lt)和肌肉的最优长度(L0)计算获得,即εt=(Lt-L0)/L0。
一个步态周期定义为脚着地到该脚再次着地。跑速定义为第4、5腰椎棘突中点在一个步态周期中的平均速度。肌肉拉伸速度为单位时间内肌肉长度的变化。采用欧拉角的方法(Kadaba et al.,1990)计算髋、膝、踝的三维角度。采用逆动力学的方法(Frankel et al.,1950)计算下肢关节的三维净力矩,其中人体惯性参数采用DeLeva(1996)修正后的Zatsiorsky-Seluyanovs人体惯性参数。关节功率为关节净力矩与关节角速度的乘积。疾跑过程中肌电的原始信号采用带通滤波进行处理,滤波器范围为10~800 Hz,之后进行全波整流,全波整流后的信号再经过20 Hz的低通滤波,获得肌电的线性包络线。采用3次样条插值法对各指标按步态周期进行归一化处理。
1.3 统计分析
采用混合设计双因素方差分析检验不同肌肉(股二头肌长头、半腱肌、半膜肌)和性别(男、女)对疾跑时肌肉最大应变及到达最大应变时间的影响,将肌肉作为重复测量因素。所有统计学分析采用SPSS 16.0软件(SPSS,Chicago,IL,USA)完成,统计学显著性的定义为一类错误概率不大于0.05。
2 结果
男性受试者的跑速为8.16±0.52 m/s,女性受试者的跑速为6.78±0.55 m/s。在跑步周期中支撑相所占百分比男、女分别为25.93%±0.03%、27.02%±0.02%。
疾跑时1个步态周期内股后肌群3块双关节肌的应变(图1)从着地开始先逐渐减小至步态周期的55%左右,之后开始增大,应变峰值出现在步态周期的90%左右(表3),此后减小至足触地。3块肌肉的收缩速度在应变减小阶段表现为负值,在应变增大阶段表现为正值(图1)。

方差分析结果显示(表2),疾跑过程中股后肌群不同肌肉的最大应变有显著性差异(F =6.156,P =0.003),但在性别间差异不显著(F =0.757,P =0.387),肌肉与性别的交互作用不显著(F =0.682,P =0.507)。后续检验表明,股二头肌长头(P =0.002)和半腱肌(P =0.029)的最大应变大于半膜肌。
肌肉与性别对疾跑过程中股后肌群最大应变出现时间(表3)的影响有显著的交互作用(F =8.213,P =0.001)。后续独立样本t检验表明,股二头肌长头(F =5.976,P =0.515)、半膜肌(F =7.480,P =0.657)和半腱肌(F =8.904,P =0.084)最大应变出现时间在性别之间无显著性差异。单因素重复测量方差分析结果表明,不同肌肉最大应变出现时间在男(F =50.041,P =0.001)、女(F =99.598,P =0.001)上均有显著性差异,后续检验表明男性(P=0.001,P =0.001,P =0.001)与女性(P =0.001,P =0.001,P =0.018)均呈现半膜肌最大应变出现时间晚于股二头肌长头和半腱肌,股二头肌长头晚于半腱肌。
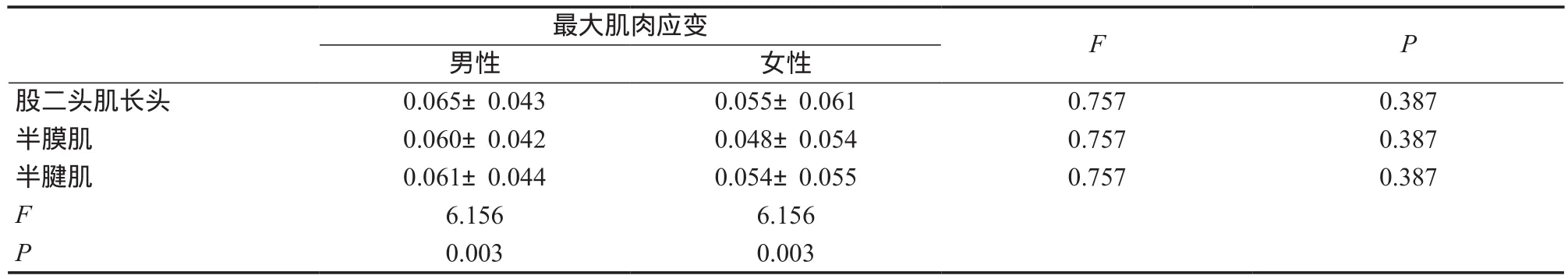
表2 疾跑过程中股后肌群的最大肌肉应变Table 2 Peak Muscle Strains during the Sprinting Gait Cycle
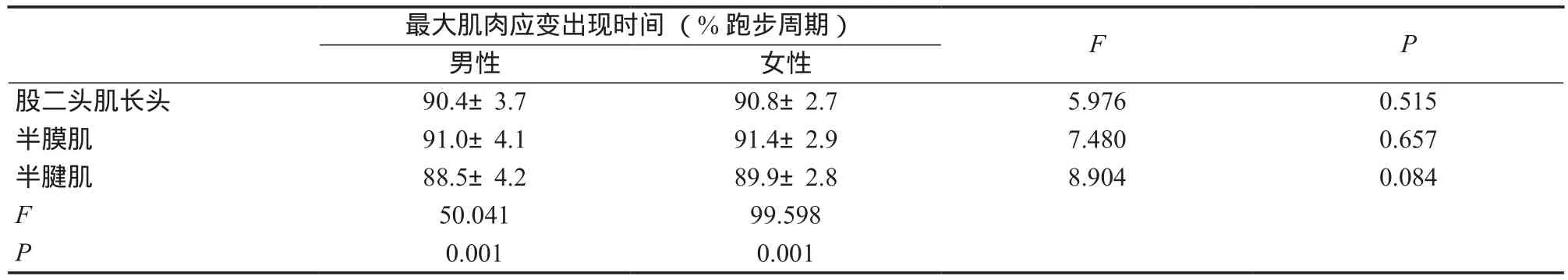
表3 疾跑过程中股后肌群最大肌肉应变出现时间Table 3 Time of Occurrence of Peak Muscle Strain during the Sprinting Gait Cycle
下肢髋、膝、踝关节矢状面关节净力矩(图2)表明,在触地初期,髋、膝、踝关节表现为伸髋、屈膝和跖屈力矩,之后的支撑相主要表现为伸髋、伸膝和跖屈力矩。在摆动相,髋关节先后表现为屈髋和伸髋力矩,而膝关节则先后表现为伸膝和屈膝力矩,摆动相踝关节处于放松,力矩几乎为零。矢状面关节功率曲线(图2)表明,在支撑相前期,伸髋力矩做正功,支撑相末期至摆动相初期,屈髋力矩做负功,之后屈髋力矩做正功,摆动相中后期伸髋力矩先做负功后做正功。支撑相膝关节主要为伸膝力矩先做负功后做正功,摆动相前期伸膝力矩做负功,摆动相后期屈膝力矩做负功。踝关节在支撑相表现为跖屈力矩先做负功后做正功,摆动相功率几乎为零。

股后肌群的肌电线性包络线的变化(图3)表明,内外侧的股后肌群均在疾跑支撑相初期和摆动相末期激活程度较高,而在支撑相末期和摆动相初、中期激活程度较低。

图3 疾跑步态周期内股后肌群的肌电线性包络线Figure 3. Linear Envelop EMG of Hamstring during the Sprinting Gait Cycle
3 讨论
疾跑过程中股后肌群拉伤机制的一个重要问题便是明确拉伤易发生的时相。本研究的结果表明,股后肌群3块双关节肌的最大应变均出现在疾跑摆动相末期,此前有学者发现,在跑台上(Thelen et al.,2005)和地面上(Wood,1987;Yu et al.,2008)跑时,股后肌群的最大长度出现在摆动相末期。研究已经表明,肌肉拉伤是离心收缩状态下肌肉应变过大导致的。本研究的结果和前人的研究共同表明,疾跑摆动相末期是股后肌群拉伤的危险时相。此外,有研究(Wood,1987;Yu et al.,2008)还发现,在疾跑支撑相末期,股后肌群也存在一个长度峰值,因此,他们认为,支撑相末期也是一个危险时相。这种前人研究之间的差异可能是跑台上跑和地面上跑之间动作模式不同导致的(Yu et al.,2008)。而本研究的结果并不支持支撑相末期是危险时相的结论,对比本研究和Yu等(2008)的研究方法可以发现,他们研究中的受试者在疾跑时可能仍处在明显的加速阶段,因为受试者的起跑位置距离动作采集区域仅有约10 m。有研究(Higashihara et al.,2015)表明,与疾跑速度保持阶段躯干基本保持直立相比,在加速跑阶段躯干倾向于前倾姿势,躯干前倾会使得骨盆前倾角和髋关节屈角增大,骨盆前倾和髋关节屈角增大使得股后肌群被拉得更长,而这便可能导致在疾跑加速跑的支撑相后期股后肌群存在一个长度峰值。
部分研究者根据屈膝和伸髋力矩峰值出现时间认为,支撑相初期(Mann,1981;Sun et al.,2015;魏书涛 等,2009)和摆动相末期(Sun et al.,2015)是疾跑股后肌群拉伤的高危时刻。本研究的下肢关节力矩结果与前人的发现基本一致,在着地初期,由于地面反作用力通过髋、膝关节的前方,地面反作用力在髋、膝关节处产生了一个很大的屈髋、伸膝力矩,为了抵抗地面反作用力的作用,腿部的肌肉,主要是股后肌群收缩产生了较大的伸髋、屈膝力矩。在摆动末期,股后肌群收缩产生一个很大的屈膝、伸髋力矩来抵抗小腿加速前摆。从肌电信号也可以发现,股后肌群在支撑相初期和摆动相末期处于剧烈收缩的状态。但是,从本研究的结果可以发现,股后肌群的肌肉拉伸速度在疾跑支撑相为负值,即处于缩短的状态。在支撑相初期,下肢髋、膝关节的净功率为正值,伸髋屈膝肌群做正功,意味着肌肉向心收缩产生能量并向肢体传递。上述结果均表明,在疾跑支撑相初期股后肌群是处于向心收缩状态的,尽管这一阶段肌肉会产生巨大的屈膝和伸髋力矩,但肌肉拉伤是由过大的应变导致的,而巨大的肌肉收缩力并不是必要条件,目前没有研究可以直接表明肌肉会在向心收缩状态下发生拉伤。因此,本研究并不支持疾跑支撑相初期是股后肌群易拉伤的危险时相。
研究结果显示,男女在疾跑时的股后肌群最大应变并没有显著性差异,这一结果提示,疾跑时股后肌群的拉伤风险可能没有性别差异。然而,这与许多临床研究的结论并不一致,很多研究发现,在各种涉及高速奔跑的运动中,男性的股后肌群拉伤率均高于女性(Dalton et al.,2015;Opar et al.,2014)。造成这一结论不同的原因可能是在比赛和训练中,男女之间的运动强度和疲劳程度不同。许多研究已经表明,疲劳是股后肌群拉伤的一个危险因素,大部分的股后肌群拉伤发生都在比赛或训练的后程(Brooks et al.,2006;Woods et al.,2004)。疲劳的肌肉抵抗被拉长的能力下降,在吸收相同能量的情况下,相比未疲劳的肌肉,疲劳的肌肉会被拉得更长,从而更容易达到被拉伤的应变条件(Mair et al.,1996)。疲劳程度的增加使得人体的本体感觉能力下降(Allen et al.,2010),改变人疾跑时的运动学和动力学特征,从而增加股后肌群的长度(Small et al.,2009,2010),进而影响肌肉的最大应变。今后的研究需要进一步确定性别和疲劳对疾跑时最大应变的影响,从而明确性别和疲劳是否对最大应变的影响产生交互作用,最终有助于理解性别对股后肌群拉伤风险的影响机制。
研究结果显示,股二头肌长头和半腱肌的最大应变大于半膜肌,这一结果提示,在疾跑时,股二头肌长头和半腱肌比半膜肌的拉伤风险更高。这与许多临床研究的结果一致,本研究的结果可以部分解释流行病学呈现出的现象。流行病学的研究结果表明,股二头肌长头是股后肌群多块肌肉中拉伤率最高的肌肉(Askling et al.,2007;Slavotinek et al.,2002),一些研究表明,半腱肌的拉伤率高于半膜肌(De Smet et al.,2000;Slavotinek et al.,2002),但也有部分研究显示半膜肌的损伤率高于半腱肌(Koulouris et al.,2003;Speer et al.,1993)。此外,本研究结果还显示,股二头肌长头和半膜肌疾跑时最大肌肉应变出现时间晚于半腱肌,半膜肌晚于股二头肌长头,表明,不同股后肌群肌肉拉伤出现的时刻可能不同。但是,通过现有的技术,临床上要获得股后肌群不同肌肉拉伤的准确时刻还很困难,今后还需要进一步验证本研究的结果。
4 结论
疾跑时股后肌群最大应变出现在摆动相末期,提示,股后肌群拉伤更易发生于疾跑摆动末期。疾跑时股后肌群最大应变无性别差异。疾跑时股后肌群各肌肉最大应变不同,这可能是疾跑时各肌肉拉伤率不同的原因之一。
