干旱和复水对两种葡萄砧木叶片光合和叶绿素荧光特性的影响
2019-03-05李敏敏袁军伟刘长江尹勇刚郭紫娟赵胜建
李敏敏,袁军伟,韩 斌,刘长江,孙 艳,尹勇刚,贾 楠,郭紫娟,赵胜建
(河北省农林科学院昌黎果树研究所,河北 昌黎 066600)
随着水资源问题日益突出,全球性干旱将日趋明显,水资源短缺也将会持续加剧[1],有关植物干旱适应性亟需进一步加强研究[2-3]。干旱引起植物体一系列生理生态及形态上的响应,导致植物光合作用降低,从而影响植物生长和产量[4-6],干旱胁迫下植物光合系统的损伤及修复对植物光合作用影响仍需进一步研究。以往有关植物干旱胁迫生理生态变化的研究大量集中在干旱胁迫期间植物的生理生化响应[7-10],但胁迫解除以后对植物的后续生长将产生怎样的影响, 胁迫期间对植物造成的不利影响能否随胁迫的解除而消除等,胁迫后复水条件下植物的生理响应研究较少[10-11],胁迫后恢复生长能力的大小通常被认为是评价植物是否抵抗逆境的重要依据[12]。
光合作用是植物生长最重要的生理过程, 对干旱胁迫非常敏感,受到干旱胁迫后,小麦、葡萄、苹果等多数植物光合速率下降,严重的干旱胁迫可以导致植物叶绿体光合机构破坏[13]。叶绿素荧光动力学技术被认为是研究植物光合生理状况及植物与逆境胁迫关系的理想探针[14-15]。
葡萄(Vitisvinifera)是对干旱比较敏感的植物,前人对葡萄砧木干旱胁迫的研究主要是集中在胁迫期间地上和地下部分形态变化,细胞膜伤害和气体交换参数的影响[16-18]。干旱后复水对葡萄砧木生长的补偿效应及其光合特性的影响较少报道。本试验就两种类型砧木干旱及复水后气体交换参数、叶绿素荧光参数的变化,研究葡萄砧木干旱及复水后的光合生理,以期更加全面地探索葡萄砧木的伤害-修复-补偿机制,为葡萄砧木抗旱生理研究提供理论基础。
1 材料与方法
1.1 试验材料
通过2016-2017年砧木耐旱性试验,筛选出抗旱性极强的葡萄砧木品种1103 P(V.berlandieri×V.rupestris)和抗旱性较差的砧木品种101-14M(V.riparia×V.rupestris),本试验以上述2个砧木作为试验材料。
1.2 试验设计
试验于2017年3—8月在河北省农林科学院昌黎果树研究所创新基地进行,3月份将1年生砧木苗定植在装有基质(田园土∶沙子∶锯末=2∶1∶2)的大营养钵(下底直径15 cm,上底直径18 cm,高25 cm)中,置于避雨棚内,常规管理。待葡萄苗长出7~8片叶时,选取生长正常、一致的植株进行干旱处理。首先对试验苗浇饱和水,隔夜,然后随机取3个试验盆,称取土壤重量(W1),放烘箱中烘干至恒重,称取土壤重量(W2),根据土壤田间持水量=(W1-W2)/W2,计算试验土壤田间持水量为21.39%。
设计干旱(T,土壤田间持水量的40%~45%)及复水,对照(CK,土壤田间持水量的70%~75%)两个处理,采取每天称重补水的方法维持土壤含水量在预设范围。当干旱处理的土壤含水量下降至试验设计要求时,定义为第0天,之后每隔7 d进行一次光合速率和荧光参数测定,干旱胁迫处理第21天开始进行复水,土壤田间持水量恢复到对照组水平。
1.3 测定项目与方法
1.3.1 光合特性 选取苗木上数第6~8片功能叶,采用便携式光合仪(LI-6400XT,LI-COR公司,美国)于第0、7、14、21、28、35天上午9∶00—11∶00测定光合参数,包括叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(E)。测定控制光强为1 600 μmol·m-2·s-1。
1.3.2 叶绿素荧光参数测定 采用便携式光合仪(LI-6400XT ,LI-COR公司,美国生产)配置脉冲调制式—荧光叶室(6400-40,LI-COR公司,美国)测定各叶绿素荧光参数,将叶片充分暗适应30 min后,测定初始荧光产量(Fo)及最大荧光产量(Fm);自然光下诱导40 min后,测量光下最小荧光(Fo′)、光下最大荧光(Fm′)和稳态荧光(Fs)。其他参数计算,PSⅡ 最大光化学效率(Fv/Fm)=(Fm-Fo)/Fm,PSⅡ 实际光化学量子产量(Yield)=(Fm′-Fs)/Fm′,光化学淬灭系数(qP)=(Fm′-F)/(Fm′-Fo),非光化学淬灭系数(QPN)=(Fm-Fm′)/(Fm-Fo′),电子传递速率(ETR)=PPFD×Yield× 0.84 × 0.5。
1.4 数据处理
数据采用5个重复的平均值,用SPSS(17.0)软件采用LSD方法在P<0.05水平进行单因素显著性分析。
2 结果与分析
2.1 干旱及复水对两种葡萄砧木叶片光合参数的影响
植物光合特性中净光合速率(Pn)作为重要的参数之一,可反映植物同化CO2的能力,进而反映出植株的生长状况。由图1a可以看出,干旱胁迫下,1103P和101-14M的Pn逐渐降低,101-14M的Pn降幅显著大于1103P(P<0.05),在干旱胁迫后第0、7、14、21天,1103P的Pn分别为5.12、4.54 、4.08、3.57 μmol·m-2·s-1, 101-14M分别降为4.56、2.13、1.58、0.83 μmol·m-2·s-1。复水后,1103P和101-14M的Pn逐渐上升,但1103P的恢复能力明显强于101-14M,复水7 d后,1103P的Pn为对照的83.20%,101-14M为对照的66.31%,复水14 d后,1103P的Pn与对照无显著差异,为对照的107.30%,101-14M为对照的88.43%。
由图1b可以看出,干旱胁迫后第0、7、14、21天,1103P的气孔导度(Gs)分别为对照的66.31%、39.36%、28.39%、28.45%,在干旱胁迫0~14 d 1103P的Gs迅速降低,14~21 d变化趋于平缓;于旱胁迫后第0、7、14、21天,101-14M的Gs分别为对照的30.09%、13.21%、9.99%、10.71%,在干旱胁迫第0~7天101-14MGs迅速降低,14~21 d趋于平缓;复水后,两种葡萄砧木的Gs又迅速升高,1103P的Gs得到迅速恢复,复水第7、14天分别为对照的86.60%、104.03%,而101-14M的Gs恢复较慢,复水第7、14天分别为对照的53.82%和75.79%。
CO2为光合作用提供直接的碳源,因此胞间CO2浓度(Ci)是影响植物光合作用的重要因素。由图1c可知,在干旱后第0、7、14、21天,1103P和101-14M的Ci为对照的59.36%、55.40%、57.72%、43.86%和54.20%、46.88%、42.00%、32.21%。复水后1103P和101-14M的胞间CO2浓度逐渐升高,复水后第7、14天,1103P和101-14M的胞间CO2浓度分别为对照的84.91%、92.37%和54.31%、85.34%。
蒸腾速率(E)反映植物蒸腾作用的强弱,蒸腾作用的大小在一定程度上能反映根系对土壤水分的吸收能力。由图1d可以看出,干旱胁迫下,1103P和101-14M的E降低,在干旱后第0、7、14、21天,1103P和101-14M的E为对照的39.57%、29.72%、29.12%、23.97%和25.96%、13.13%、6.26%、3.90%。复水后,1103P和101-14M的蒸腾速率逐渐恢复,复水第7、14天分别为对照的74.54%、103.13%和55.79%、64.36%。
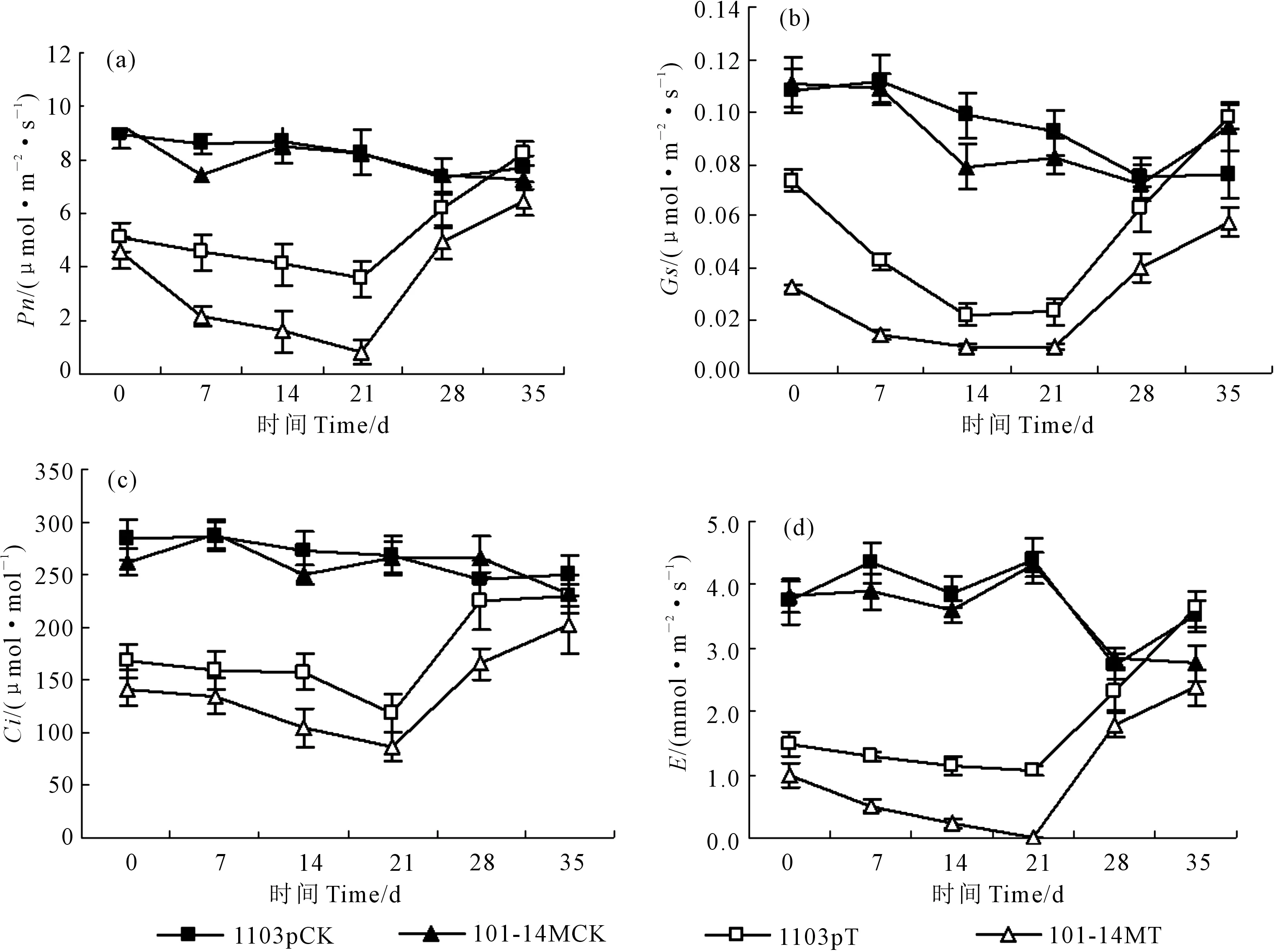
图1 干旱及复水后两种葡萄砧木光合参数的变化Fig.1 The photosynthesis changes of two grape rootstocks after drought and rewatering
2.2 干旱及复水对两种葡萄砧木叶片荧光参数的影响
干旱及复水后1103P和101-14M荧光参数变化如图2所示。暗适应下初始荧光产量(Fo)是判断PSⅡ反应中心运转的重要指标,Fo上升表明PSⅡ反应中心受到破坏或可逆失活。由图2a可知,干旱胁迫后,两种葡萄砧木的Fo都有上升趋势,1103P在干旱胁迫14 d后Fo迅速增加,而101-14M在干旱胁迫7 d后Fo迅速增加,1103P和101-14M在干旱胁迫第7、14、21天Fo分别比对照增加了0.64%、1.68%、6.28%和4.00%、13.44%、14.77%,同时期101-14M的Fo大于1103P;复水后,两种葡萄砧木的Fo逐渐降低,1103P和101-14M在复水后第7、14天分别恢复到对照的102.95%、101.56%和109.60%和101.81%。
光化学量子效率(Fv/Fm)表示PSII最大的(潜在)光化学量子效率。由图2b可知,干旱胁迫后1103P和101-14M的Fv/Fm都表现出逐渐降低的趋势,复水后逐渐增加,然而在干旱胁迫7 d时,1103P的Fv/Fm与对照无显著差异,而101-14M的Fv/Fm显著低于对照(P<0.05),复水7天后(第28天)1103P的Fv/Fm恢复到对照水平,而复水14 d后(第35天)101-14M才恢复到对照水平,干旱胁迫及复水后同时期1103P的Fv/Fm显著高于101-14M。
由图2c、2d、2e可知,干旱及复水后1103P和101-14M的Yield、qP、ETR变化趋势与Fv/Fm变化趋势基本一致,都表现为干旱胁迫后逐渐降低,而复水后逐渐增加,干旱胁迫7 d时1103P的Yield、qP、ETR与对照无显著差异,而101-14M显著低于对照(P<0.05),复水7 d(第28天)时1103P的Yield、qP恢复到对照水平,而101-14M在复水14 d(第35天)时恢复到对照水平。干旱胁迫及复水后同时期1103PYield、qP、ETR显著高于101-14M。
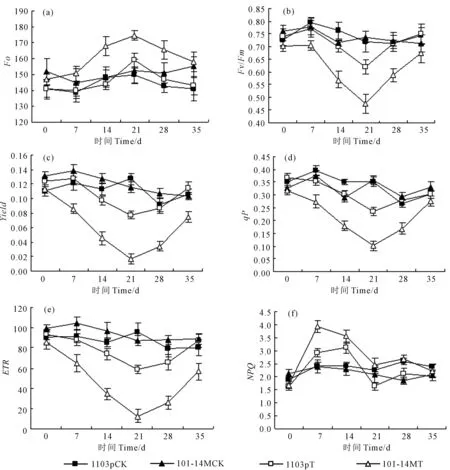
图2 干旱及复水后两种葡萄砧木叶片叶绿素荧光参数变化Fig.2 Changes of the chlorophyll fluorescence parameters of two grape rootstocks after drought stress and rewatering
非光化学淬灭系数(NPQ)反映了植物热耗散能力的变化。由图2f可知,干旱胁迫后1103P和101-14M的NPQ呈现先增加后降低的趋势,101-14M在干旱胁迫后第7天出现最大值(3.92),之后降低,说明了在干旱胁迫7 d之前,101-14M通过热耗散来散失多余热量的能力较强,之后由于受到干旱胁迫的伤害热耗散能力降低;说明1103P在第14天出现最大值(3.11),之后降低,1103P在干旱胁迫14天之后热耗散能力降低。复水后,1103P和101-14M的NPQ呈现增加的趋势,在第35天均恢复到对照水平。
3 讨论与结论
土壤干旱胁迫对植物生长和代谢的影响是多方面的,包括植株生长势、光合作用等方面,其中对光合作用的影响尤其突出和重要。通常将影响植物光合作用的因素分为气孔限制和非气孔限制,胞间CO2浓度的变化趋势与净光合速率的变化趋势比较,是判断气孔限制和非气孔限制的依据[19-20]。研究普遍认为,植物在短期逆境条件下引起光合作用降低主要是气孔限制的作用,植物在逆境胁迫下,为了降低蒸腾作用、保持水分而关闭气孔,从而使气孔导度降低,蒸腾速率降低,也引起了光合速率的降低[21-23]。本研究也印证了这一结论,在整个干旱胁迫期间,1103P和101-14M的光合速率逐渐降低,而1103P Gs在14 d之前也迅速降低,之后降低幅度明显趋于平缓,101-14M Gs在7 d之前迅速降低,之后降低幅度平缓,因此认为在干旱胁迫前期,引起光合作用降低的主要因素是气孔限制。然而在干旱胁迫后期,气孔导度不再降低,而光合速率进一步降低,根据叶片净光合速率、气孔导度、叶绿素荧光值的变化趋势,结合Medrano等的假说[24],认为随着干旱胁迫时间的延长,1103P和101-14M叶片净光合速率的降低主要是非气孔限制因素,这可能是因为光合机构的受损引起的。这一结论与前人在小冠花[25]、大豆[9]、油菜[26]等作物上的结论基本一致。
叶绿素吸收的光能,主要通过3种途径耗散:光合电子传递、叶绿素荧光和热耗散,因此叶绿素荧光在一定程度上能反映光合电子传递情况。通过叶绿素荧光参数测定,可以获得PSⅡ反应中心的状态和电子转化效率、PSⅡ电子传递活性等情况[27]。研究已证明,通过分析逆境下荧光参数的变化幅度,可以判断植物对逆境的抵抗能力[28]。在干旱胁迫后,1103P和101-14MFo出现了不同程度的增加,101-14M在干旱胁迫7 d后出现迅速增加,而1103P在14 d后出现迅速增加,而且同时期101-14MFo值显著高于1103P,说明了1103P的光合作用中心受伤害时间晚于101-14M,且同时期1103P光合反应中心的受伤害程度小于101-14M,说明了1103P的耐旱性强于101-14M的原因之一是光合反应中心的受伤害程度较低。
许多研究表明,Fv/Fm与植物的生长状态呈现高度的正相关[29]。在干旱和强光等非生物逆境胁迫下,植物吸收的光能超过自身光合作用需要的光能时,会发生光抑制,通常Fv/Fm的变化被认为是判断是否发生光抑制的标准,正常条件下Fv/Fm变化幅度较小,如果Fv/Fm明显降低,则说明发生了光抑制[20-32]。前人关于胁迫后植物是否发生光抑制现象的结论也不完全一致,棉花[32]、豌豆[33]、小麦[34]等植物在逆境条件下发生了光抑制,而在油菜[27]等植物上却未发现。本研究中,1103P和101-14M在干旱胁迫后Fv/Fm出现了不同程度降低,说明1103P和101-14M干旱胁迫后发生了光抑制。
干旱胁迫后复水可研究植物对干旱胁迫的适应能力和保护机制。一些研究表明,植物在胁迫解除后的复水条件下会出现快速生长,以弥补胁迫造成的损失,通过改善叶片的气孔运动、光合作用、气孔导度等生理代谢功能,从而使植株出现快速生长,补偿的结果主要体现在株高、叶面积、生物量等的迅速增加[9,35],有的植物甚至出现超补偿作用。本研究中,1103P和101-14M在复水后出现净光合速率迅速增加、气孔导度增加、光合系统的破坏得到恢复等,然而1103P和101-14M在复水后的恢复程度不同,1103P在复水7 d后,各项功能基本恢复到了对照水平,在复水14 d时净光合速率显著高于对照,出现超补偿作用,而101-14M在复水14 d内未出现超补偿作用。
植物的耐旱性受多种因素的影响,本试验在盆栽条件下,研究了两种不同耐旱性葡萄砧木在干旱和复水后叶片光合作用和荧光特性的变化,仅从这两方面揭示了葡萄砧木的耐旱生理机制。下一步,将从砧木叶片组织结构的差异与抗旱性的关系,以及水通道蛋白如何调节气孔的行为等方面进一步阐述葡萄砧木的耐旱机理。
