干旱胁迫对3种苹果砧木叶片光合、叶绿体超微结构和抗氧化系统的影响
2019-03-05郭爱霞石晓昀王延秀朱燕芳
郭爱霞, 石晓昀, 王延秀, 胡 亚, 朱燕芳
(甘肃农业大学园艺学院,甘肃 兰州 730070)
苹果(Maluspumila)是我国重点果产区的支柱性产业[1-2],而西北黄土高原因具备适于苹果生产的自然条件而成为我国苹果的最优生态区,栽培面积占全国苹果总种植面积的44%,产量占全国苹果总产量的49%[3],但黄土高原多为干旱或半干旱雨养农业区,水分亏缺成为制约植物生长的主要因素[4],而且果树对干旱胁迫反应敏感。水分胁迫下,果树对水分和养分的吸收减少,正常的生理活动,如光合代谢等受到干扰,体内有机物的积累和转运受滞,进而严重影响树体的生长发育和果实的产量品质[5-6]。因此,在水分供应不足的地区,筛选并应用抗旱性较强的砧木对果树的节水、优质高产具有重要意义[7-8]。
前人已经探讨了不同植物对干旱胁迫的生理响应机制,主要集中于生理生化、形态结构和基因表达的差异。干旱胁迫下植物的生理功能发生显著变化,高杰等[9]对玉米的研究结果表明,干旱胁迫下玉米干物质积累量、叶面积、株高显著下降,光系统Ⅱ和抗氧化酶系统损伤,光合速率下降。季杨等[10]针对干旱胁迫对鸭茅根及叶的保护酶活性、渗透物质含量及膜脂过氧化作用的研究表明干旱胁迫诱导2种基因型的鸭茅根系及叶片细胞产生活性氧,发生氧化反应,破坏细胞膜而降低酶活性。李泽对油桐的研究表明干旱下,SPAD、Pn、Gs、Tr、Fv/Fm及qP值降低[11]。植物细胞中的叶绿体对逆境胁迫比较敏感,干旱胁迫下叶绿体的形态、结构和大小数目均会发生变化,Zhang等[12]对甘蔗做了相关研究,发现干旱胁迫下细胞发生质壁分离,叶绿体由长椭圆变为近圆形,并向细胞中心靠近,淀粉粒变小消失。徐萍等[13]研究了干旱胁迫对银沙槐幼苗叶绿体超微结构的影响,结果表明干旱胁迫下银沙槐细胞的叶绿体变形,外被膜波浪状、断裂不完整;基粒和基质类囊体膜结构模糊,基粒弯曲膨胀。Rampino[14]研究了普通小麦和山羊草在分子水平上响应干旱胁迫的差异,结果发现通过RT-PCR分析的DHN基因在供水充足的小麦中不表达,而在受胁迫的植物中表达。Yoshimura利用差异蛋白质组学研究认为,旱生植物野生西瓜在胁迫过程中由避旱性转变为耐旱性,通过时间程序化的方式调节它的根蛋白质组[15]。
目前对苹果属植物的耐旱性研究较多[16-17],但多数仅从生理特性或解剖结构单方面进行抗性比较,而结合PSII光合系统、抗氧化系统和叶绿体超微结构的变化来综合评价苹果砧木抗旱性的研究未见报道。本试验以新疆野苹果(MalussieversiiRoem.(XJ))、垂丝海棠(MalushallianaKoehne(CS))及山定子(MalusbaccataBorkh.(SDZ))为研究对象,初步探讨了这三种苹果砧木的干旱适应机制,运用相关性分析和主成分分析法综合评价它们的抗旱性,旨在为苹果抗旱砧木的选育提供理论依据。
1 材料与方法
1.1 试验材料及处理
试验于2016年4—6月在甘肃农业大学园艺学院避雨棚进行。选取生长一致的新疆野苹果(XJ)、垂丝海棠(CS)及山定子(SDZ)一年生幼苗,移入盛装基质(20%蛭石,20%珍珠岩,60%泥炭)重量为0.65 kg的花盆(内径10 cm,深18 cm)中,每盆1株,统一管理30 d后,进行盆栽控水胁迫,分为正常供水(土壤含水量为田间持水量的75%~80%)和干旱胁迫(土壤含水量为田间持水量的45%~50%)两个处理,每个处理30盆幼苗。采用持续称重法控制土壤含水量,每隔3 d傍晚称重并补充水分。当最大田间持水量为100%时,土壤净含水量为48%。试验所设处理分别为新疆野苹果正常供水(XJ75)及干旱胁迫(XJ50)苗木、垂丝海棠正常供水(CS75)及干旱胁迫(CS50)苗木、山定子正常供水(SDZ75)及干旱胁迫(SDZ50)苗木,从胁迫前1天(记为第0天)开始,每隔7 d测定一次各项指标,重复5次。
1.2 测定项目与方法
1.2.1 光合参数的测定 从胁迫处理0 d开始,每隔7 d上午9∶00选取生长一致的幼苗相同节位的叶片,将各项指标逐一测定,重复5次。用Li-6400光合仪(LI-COR公司,美国)测定第8片真叶的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)。设定CO2浓度为400 μmol·mol-1,环境温度25℃,光强为800 μmol·m-2·s-1。
1.2.2 叶绿素荧光参数的测定 在暗适应后,设置测量光饱和闪光后夹好叶片,用Li-6400光合仪(LI-COR公司,美国)测定荧光参数,等待dF/dt绝对值<5时记录数据,计算PSII最大光化学效率(Fv/Fm)、PSII潜在光化学活性(Fv/Fo)、光化学猝灭系数(qP)、非光化学猝灭系数(NPQ)等光合荧光参数,重复测定5次。
1.2.3 透射电镜切片的制备 干旱胁迫28 d后,取对照及处理植株新梢顶端(形态学顶端)完全展开的第4片幼叶,用去离子水冲洗干净后,将叶片剪成1 mm×1 mm的组织块,立即投入用0.1 mol·L-1磷酸缓冲液(PBS,pH7.4)配制的3%戊二醛中抽气5~8 min,组织块下沉后,用PBS配制的3%戊二醛于4℃下固定12 h,PBS冲洗3次,每次15 min;用PBS配制的1%锇酸在4℃下固定2 h,用PBS冲洗3次,每次15 min;乙醇梯度脱水,依次用50%、70%、80%、90%、100%的乙醇浸泡,每梯度脱水15 min,再用纯丙酮脱水2次,每次15 min;脱水后进行渗透与包埋,100%丙酮∶包埋剂=1∶1,渗透5 h,然后换成纯包埋剂包埋16 h(包埋剂由9.12 mL环氧树脂Epon812+4.7 mL 软化剂MNA+6.15 mL硬化剂DDSA+0.3 mL催化剂DMP-30 配制而成),最后在36℃下聚合12 h,72℃下聚合48 h。
用LEICA EM UC6 Miultracut超薄切片机(Leica Microsystems GmbH,Wetzlar,Germany)切出厚度约为70 nm的超薄切片,用醋酸双氧铀和柠檬酸铅双重染色后,用TECNAI G2(TF20)型透射电镜(FEI,USA)对叶片叶绿体超微结构进行观测,超薄切片每种植物每个处理观察20个视野并照相。
1.2.4 抗氧化酶活性的测定 使用高锰酸钾滴定法[18]测定过氧化氢酶(CAT)活性;NBT光还原法[19]测定超氧化物歧化酶(SOD)活性;愈创木酚法[20]测定过氧化物酶(POD)活性。
1.3 数据处理
用Excel 2013进行数据统计整理,使用软件origin 9.0作图,采用R语言软件进行相关性分析,采用SPSS 22.0进行主成分分析。
2 结果与分析
2.1 干旱胁迫对三种苹果砧木叶片光合特性的影响
如图1A所示,与正常供水处理相比,干旱胁迫下, 3种苹果砧木的净光合速率(Pn)降低,随着胁迫时间的持续,XJ、CS和SDZ的Pn均呈下降趋势,胁迫28 d后,它们的Pn分别降低了35.20%、12.75%、43.13%,表明CS的Pn受干旱胁迫的影响较小。3种苹果砧木的气孔导度(Gs)变化趋势基本与Pn的变化趋势相似(图1B),干旱胁迫下XJ、CS和SDZ的Gs显著下降,随着胁迫时间的持续,CS的气孔导度下降了7.10%,降幅小于XJ和SDZ。 干旱胁迫下,3种苹果砧木的Tr均显著降低(图1C),胁迫处理28 d后,XJ、CS和SDZ的Tr降幅分别为:29.40%、16.58%、35.94%,CS的降幅小于XJ和SDZ,说明CS受干旱胁迫的影响程度要小于XJ和SDZ。如图1D,相比正常供水处理,干旱胁迫下胞间CO2浓度(Ci)的变化趋势与前3个光合参数变化趋势相反,随着干旱胁迫的持续,XJ、CS和SDZ的Ci均呈现上升趋势,其中CS的增幅较大。
2.2 干旱胁迫对3种苹果砧木叶片光合荧光特性的影响
从图2A可知,干旱胁迫期间,XJ、CS和SDZ的最大光化学效率(Fv/Fm)与对照相比均呈现下降趋势,降幅分别为:13.32%、7.04%、14.23%,其中CS的降幅低于XJ和SDZ。XJ、CS和SDZ潜在光化学活性(Fv/Fo)的变化趋势与Fv/Fm相似(图2B),干旱胁迫显著降低了3种苹果砧木的Fv/Fo,胁迫处理28 d后,CS的降幅较XJ和SDZ小。如图2C所示,随着干旱胁迫时间的延长,3种苹果砧木的光化学猝灭系数(qP)均呈下降趋势,XJ的qP下降了18.17%,CS和SDZ的qP分别下降了5.96%和22.01%,CS的降低程度明显低于XJ和SDZ。随着干旱胁迫的持续,3种砧木叶片的非光化学猝灭系数(NPQ)均呈上升趋势(图2D),在胁迫28 d后,增幅分别为12.78%、20.74%、7.37%,CS的NPQ增幅较XJ和SDZ的大。
2.3 干旱胁迫对3种苹果砧木叶片叶绿体超微结构的影响
正常供水条件下,3种砧木的叶肉细胞具有清晰、光滑、连续的细胞膜和细胞壁,叶绿体均为椭圆形,并紧贴细胞壁分布,每个叶绿体中具有1~2个淀粉颗粒,叶肉细胞内有一个大的液泡(图3A、C、E);核仁和染色质在核基质中分散(图3A);基质中含有线粒体和许多质体小球及较大淀粉粒的组织良好的叶绿体(图3A、C)。干旱胁迫下,3种砧木叶肉细胞表现出明显的结构变化,出现明显的质壁分离现象,淀粉粒缩小甚至消失(图3B、F);染色质和核仁缩合甚至消失(图3B);叶绿体肿胀,但淀粉颗粒和线粒体基本无变化(图3D);叶绿体发生变形和空泡化,线粒体和质体小球消失(图3F)。3种砧木的叶绿体超微结构受到不同程度的伤害,但相比其他两种砧木,CS的叶片超微结构损伤较小,能较好地保持细胞结构的完整性。
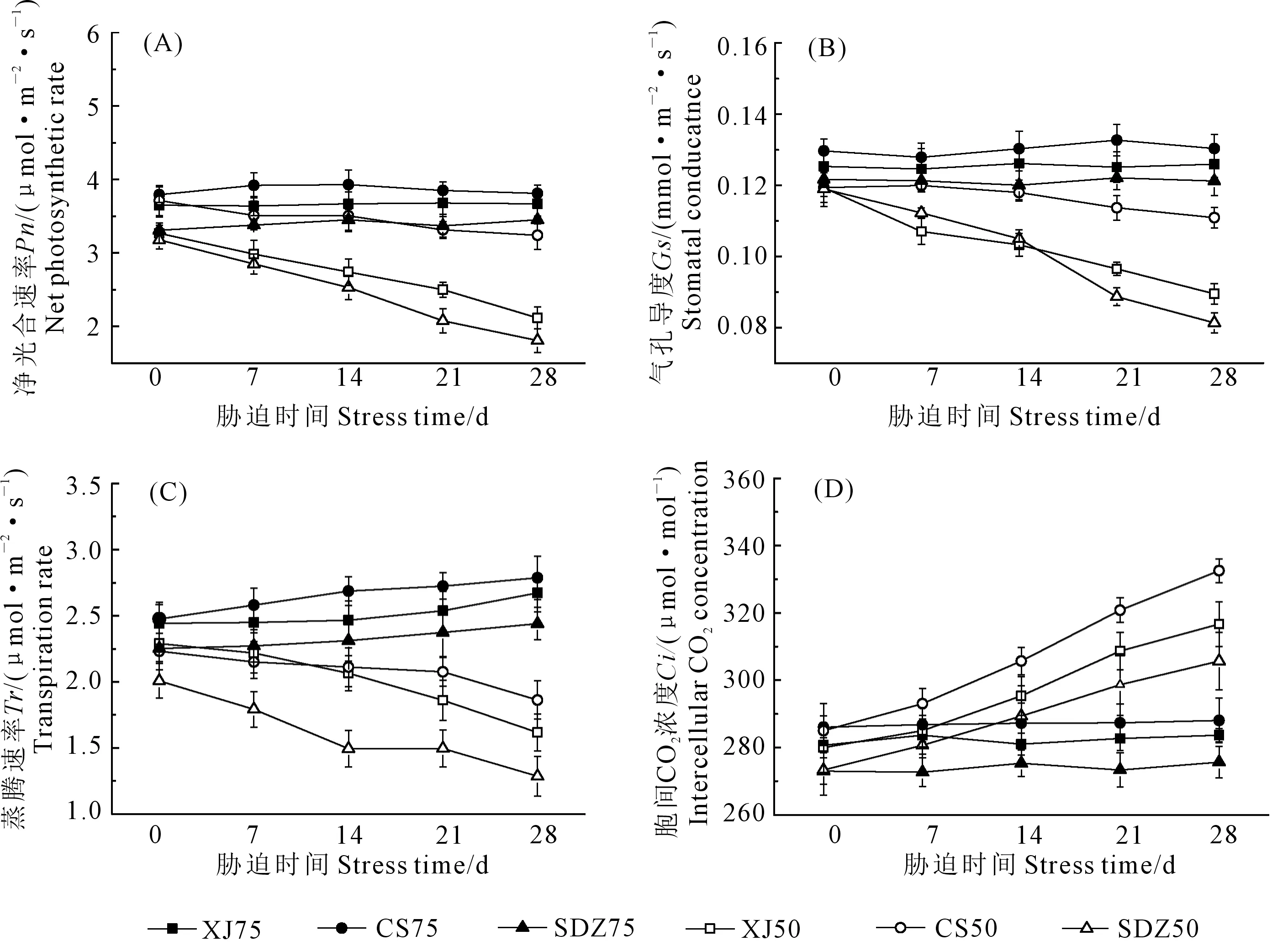
注:XJ75-新疆野苹果正常供水;CS75-垂丝海棠正常供水;SDZ75-山定子正常供水;XJ50-新疆野苹果干旱胁迫;CS50-垂丝海棠干旱胁迫;SDZ50-山定子干旱胁迫。下同。Note: XJ75 represents adequate water supply condition of Malus sieversii Roem; CS75 represents adequate water supply condition of Malus halliana Koehne; SDZ75 represents adequate water supply condition of Malus baccata Borkh; XJ50 represents drought stress of Malus sieversii Roem; CS50 represents drought stress of Malus halliana Koehne; SDZ50 represents drought stress of Malus baccata Borkh. The same below.图1 干旱胁迫对3种苹果砧木叶片光合特性的影响Fig.1 Effects of drought stress on photosynthetic characteristics of leaves of three apple rootstocks
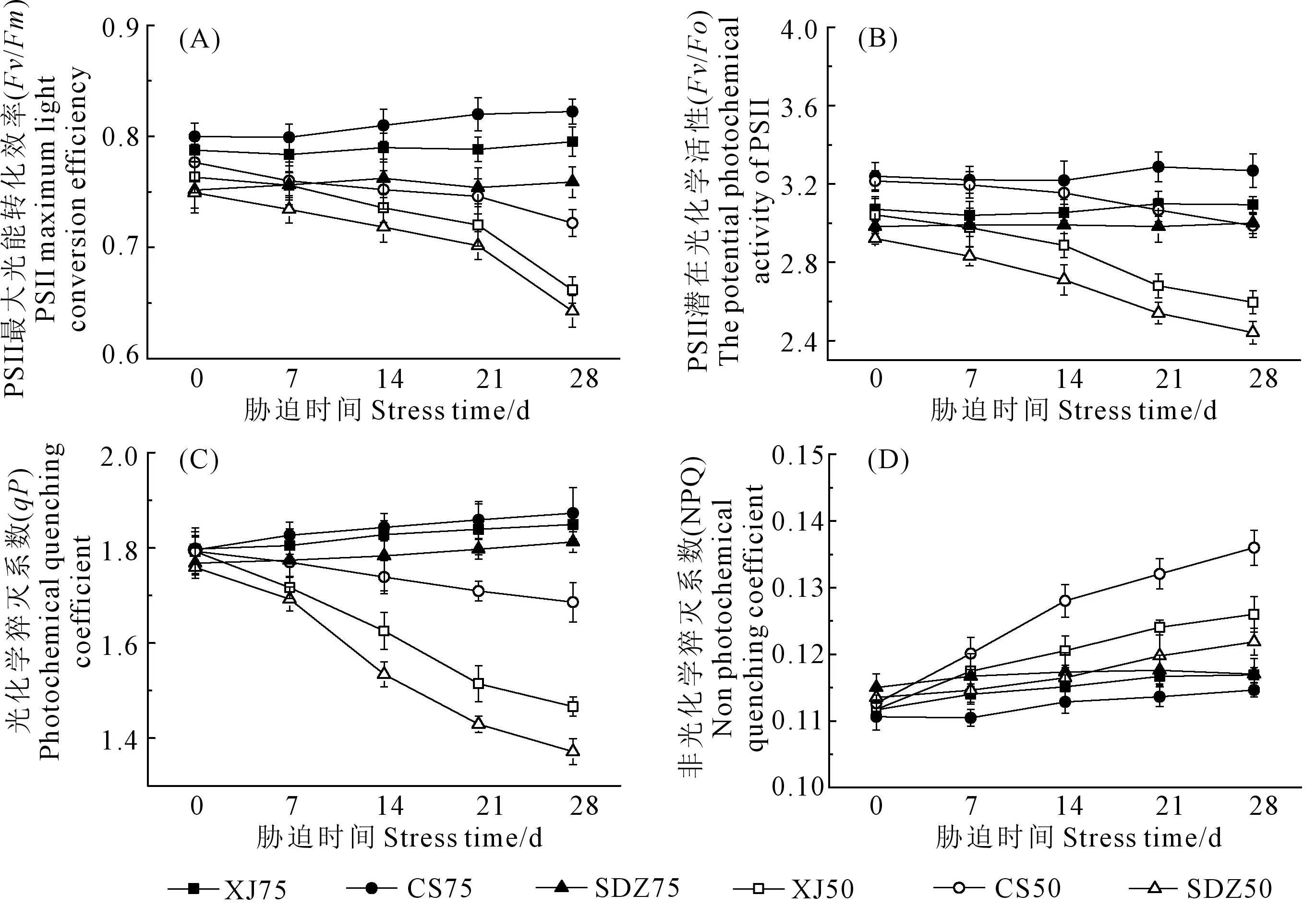
图2 干旱胁迫对3种苹果砧木叶片荧光特性的影响Fig.2 Effects of drought stress on fluorescence characteristics of leaves of three apple rootstocks
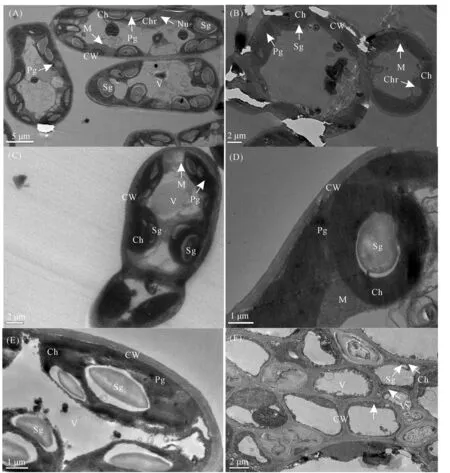
注:A、C、E为正常供水条件下的XJ、CS、SDZ,B、D、 F为水分胁迫下的XJ、CS、SDZ。CW:细胞壁;Ch:叶绿体;Sg:淀粉颗粒;Chr:染色质;Pg:质体小球;Nu:核仁;M:线粒体;V:液泡;Vs:基质片层囊泡化。Note:A, C, and E denote XJ, CS, and SDZ under control conditions, respectively. B, D, and F denote XJ, CS, and SDZ under drought conditions, respectively. CW: cell wall; Ch: chloroplast; Sg: starch grain; Chr: chromatin; Pg: plastoglobule; Nu: nucleolus; M: mitochondrion; V: vacuole; Vs: vesiculation of stroma lamellae.图3 干旱胁迫对3种苹果砧木叶片叶绿体超微结构的影响Fig.3 Effects of drought stress on chloroplast ultrastructure of leaves of three apple rootstocks
2.4 干旱胁迫对3种苹果砧木叶片抗氧化系统的影响
2.4.2 干旱胁迫对3种苹果砧木叶片抗氧化酶活性的影响 干旱胁迫下,CS、XJ以及SDZ的SOD和CAT活性均呈先上升后下降的变化趋势(图5A和图5C),胁迫处理14 d时,它们的SOD和CAT活性均达到最大值,整体呈升高趋势;从图5B可知,胁迫处理前21 d,3种砧木的POD活性呈现升高趋势,21 d之后趋于平稳。总体来讲,CS的SOD、POD和CAT活性增幅高于XJ和SDZ。
2.5 3种苹果砧木叶片各生理指标的相关性分析
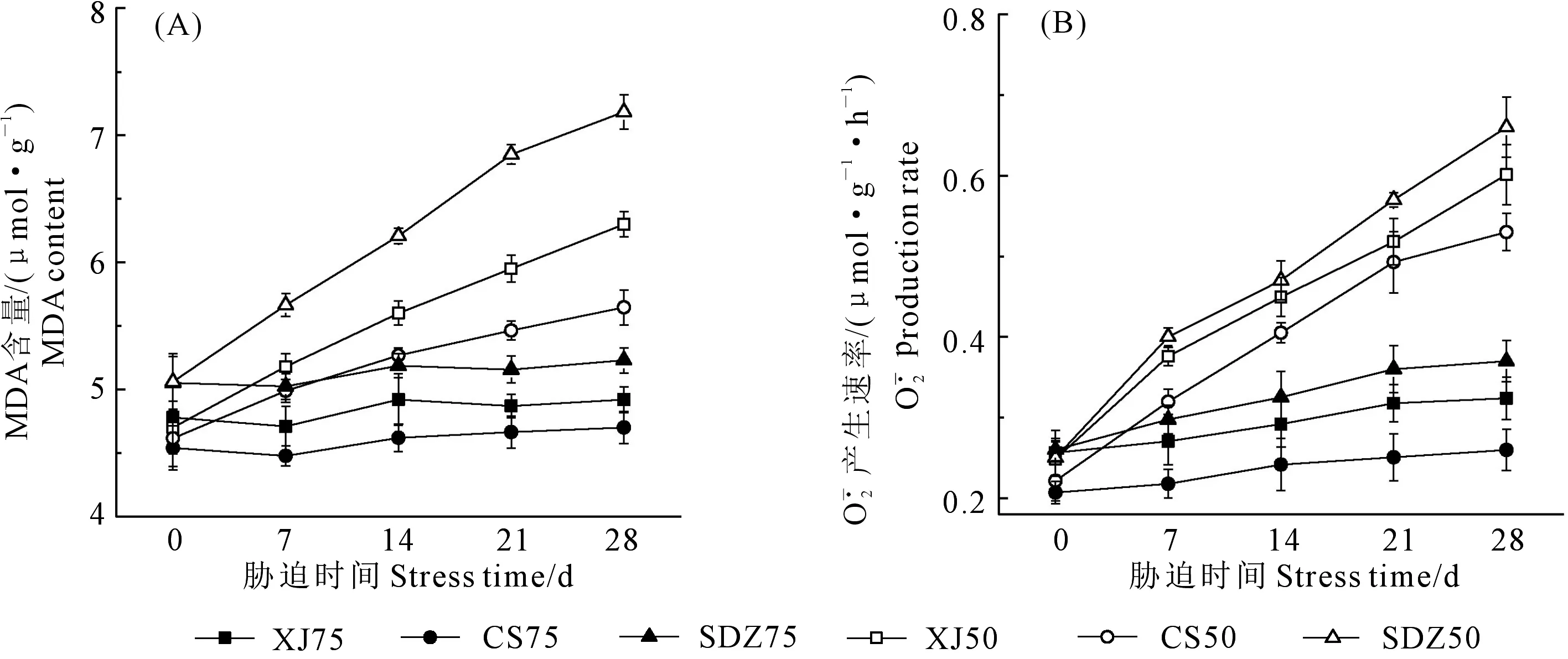
图4 干旱胁迫对3种苹果砧木叶片MDA含量和产生速率的影响Fig.4 Effects of drought stress on MDA content and production rate of leaves of three apple rootstocks
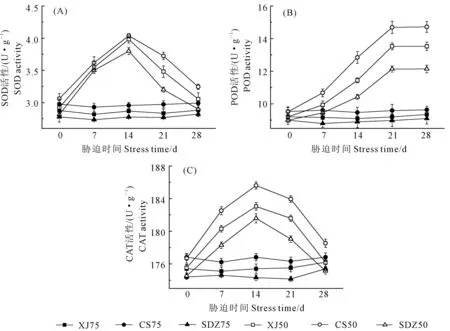
图5 干旱胁迫对3种苹果砧木叶片抗氧化酶活性的影响Fig.5 Effects of drought stress on activities of three kinds of antioxidant enzymes of leaves of three apple rootstocks
2.6 3种苹果砧木叶片各生理指标之间的主成分分析

3 讨 论
3.1 干旱胁迫下3种苹果砧木叶片光合特性的分析
干旱胁迫期间,植株的正常光合作用受到气孔和非气孔限制因素的影响,Farquhar 和Sharkey[21]研究发现,引起光合速率降低的气孔或非气孔限制因素可以根据Ci的变化方向来判断,当光合作用和Ci均降低时,光合作用受到了气孔限制因素的影响,相反,Ci升高则光合作用受到非气孔限制的影响。也有研究[22]表明水分胁迫下苹果幼苗叶片的Pn、Gs和Ci均显著降低。本研究中,3种苹果砧木的Pn、Gs和Tr随着干旱胁迫的持续均呈下降趋势,而Ci则呈现升高趋势,所以干旱胁迫下造成苹果叶片光合作用下降的因素是非气孔限制,而非气孔限制因素是由于光合细胞机构和功能受到了损伤导致的。一年生的3种砧木幼苗,可能是由于长期培养于避雨棚中,受光条件较弱,苹果叶片质地较薄,因此Pn值均偏小。而CS在干旱胁迫期间,Pn、Gs、Tr和Ci的变化幅度显著低于XJ与SDZ,可能是由于其叶片利用荧光或热能耗散的形式,阻止过多光能向PSII光系统中心传递,以保护光合作用机构免于强光破坏,有利于光合作用正常进行,这与Singh的研究结果相类似[23]。

注:图中黑色圆圈表示两者呈正相关,白色圆圈表示两者呈负相关,圆圈越大,相关性越高。Note: The black circles in the figure indicate a positive correlation between the two indices, the white circle indicate a negative correlation between the two indices. The larger the circles, the higher the correlation.图6 3种苹果砧木叶片各生理指标间的相关系数Fig.6 The correlation coefficient between the physiological indexes of leaves of the three apple rootstocks
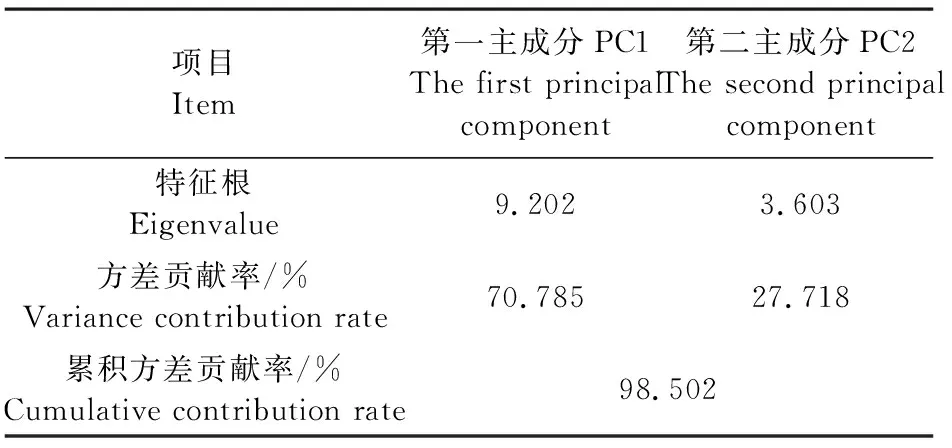
项目Item第一主成分PC1The first principalcomponent第二主成分PC2The second principalcomponent特征根Eigenvalue9.2023.603方差贡献率/%Variance contribution rate70.78527.718累积方差贡献率/%Cumulative contribution rate98.502
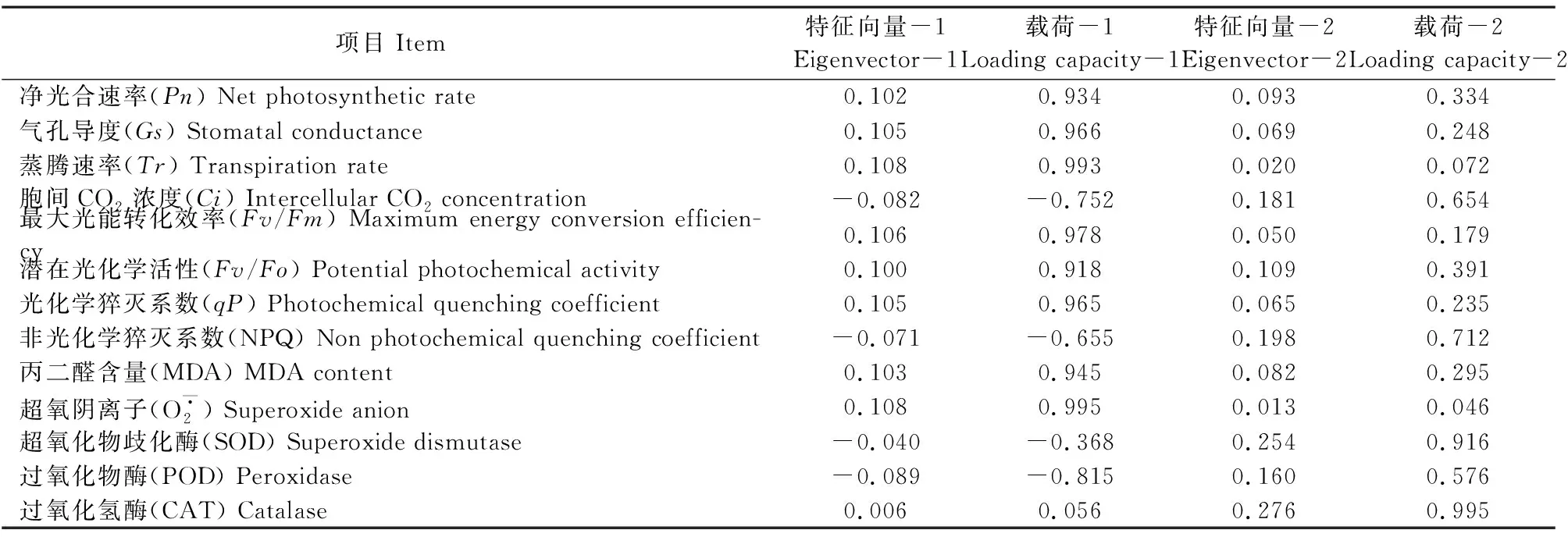
表2 各主成分的特征向量
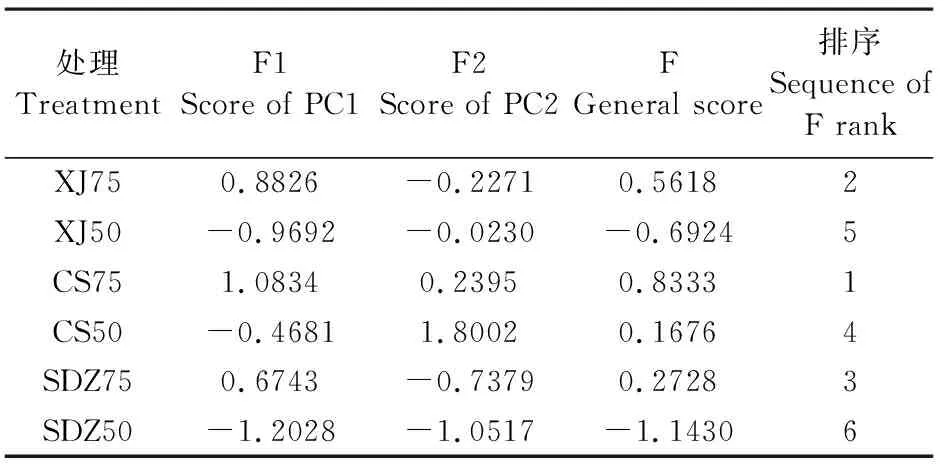
表3 各主成分得分及综合得分
叶绿素荧光参数是干旱胁迫期间量化植物水分状况的一种工具,能够快速、准确、无伤害地反映干旱胁迫对光合作用产生的影响[24]。Fv/Fm和Fv/Fo是反映PSII反应中心光合潜能的重要参数;qP反映PSII天线色素捕获的光能用于光化学电子传递的份额,其下降幅度越小,表明越抗旱;NPQ反映PSII天线色素吸收的光能以热的形式耗散掉的部分,较高的NPQ有利于过剩光能的及时耗散,从而避免光系统的损害[25]。干旱胁迫下,PSII光系统通过提高非辐射性热耗散,消耗PSII吸收的过剩光能,从而保护PSII反应中心免受因吸收过多光能而造成的光氧化伤害[26]。从本研究结果可知,随着干旱胁迫时间的延长,XJ、CS和SDZ的Fv/Fm、Fv/Fo和qP均降低,NPQ上升,表明干旱胁迫导致三种砧木光合电子传递受阻,类囊体膜受到损伤引起光合速率下降,与其它两种砧木相比,CS的Fv/Fm、Fv/Fo和qP降幅较低,NPQ的上升程度较高,表明干旱胁迫下CS具有较强的热耗散和电子传递能力,PSII中心光能转化效率较高,具有较高的光能利用率,对干旱胁迫的适应能力强,这与牛铁泉[22]等的研究结果相一致。
3.2 干旱胁迫下3种苹果砧木叶片超微结构的分析
正常供水条件下,叶肉细胞中的叶绿体都紧贴于细胞壁,这种分布方式有利于大气中的CO2向叶绿体中扩散,促进光合作用[27]。干旱胁迫下,细胞液泡收缩,核染色质凝聚,细胞发生质壁分离,叶绿体膜结构被破坏,淀粉粒变小消失,叶绿体弯曲膨胀,基粒类囊体空泡化[17]。本试验结果发现,与XJ和SDZ相比,干旱胁迫期间CS的叶绿体受伤害程度较轻。叶绿体是生成活性氧(ROS)的主要来源,所以易受氧化损伤的影响,对干旱胁迫反应较敏感,而超微结构的损伤程度间接取决于调节ROS水平的抗氧化酶浓度的变化,由于CS的SOD、POD和CAT水平高于XJ和SDZ,干旱胁迫期间酶活性降幅也较低,所以CS的膜脂过氧化程度较低,叶绿体细胞膜受损较轻,与前人研究一致[17]。
3.3 干旱胁迫下3种苹果砧木叶片抗氧化系统的分析
主成分分析(PCA)方法将原来众多具有一定相关性的多个指标重新组合成一组新的互相无关的综合指标,消除了数据之间的相关性[34],操作简单,没有参数限制,可应用于多方面研究[35]。为了全面综合地比较干旱胁迫下3种苹果砧木的抗旱性,本试验采用PCA对3种砧木的所测指标进行评价排序,将原来的13个指标简化为2个主成分,对这2个主成分的载荷进行分析发现,干旱胁迫下CS的PCA得分高于XJ和SDZ,说明CS的抗旱性较强。
4 结 论
对干旱地区常用苹果砧木新疆野苹果、垂丝海棠和山定子进行抗旱性评价发现,垂丝海棠能在干旱胁迫下保持叶绿体结构的完整性,激活抗氧化酶系统,清除氧化产物而保持较高的光合能力。根据PCA综合分析得出,垂丝海棠的抗旱性强于新疆野苹果和山定子。
