miR-199a-3p靶向视网膜母细胞瘤转录辅阻遏子1促进心肌细胞肥大
2019-03-01胡志琴朱杰宁符永恒袁淑菁张梦珍单志新
杨 静,胡志琴,朱杰宁,李 晖,符永恒,袁淑菁,潘 蓉,张梦珍,单志新,
(1.华南理工大学医学院,广东广州 510006;2.广东省人民医院//广东省医学科学院//广东省心血管病研究所,广东广州510080;3.南方医科大学药学院,广东广州 510515)
在成人心脏中,心脏增大不是心肌细胞数量增加,而是单个心肌细胞的大小增加。心肌细胞肥大最初是对生理和病理刺激的适应性反应,但是病理性肥大通常发展为心力衰竭。由于病理性刺激(如压力超负荷)引起的进行性心脏肥大常与心力衰竭的发展相关,心力衰竭是全球发病率和死亡率的主要原因[1]。研究表明,以前未被认识的机制,包括细胞代谢,增殖,非编码RNA,免疫反应,翻译调控和表观遗传修饰,都会正向或负向调节心脏肥大[2]。microRNA(miRNA)是进化上保守的非编码RNA,长约22个核苷酸,其通过翻译抑制或降解靶mRNA从而负向调节基因表达[3-4]。越来越多的证据表明miRNA广泛参与心脏肥大的发病机制[4-7]。MiR-1通过靶向抑制许多信号分子,包括真核起始因子4E(Eif4e),Mef2a、Gata4、组蛋白脱乙酰酶6(HDAC6)和CDK6的翻译来保护心脏结构和功能以抑制心肌肥厚[8-11]。MiR-133a通过调节Akt及其下游信号分子如Cdc42、Rho-A和Nelf-A/WHSC2来减弱心肌肥厚[12]。MiR-10a和miR-497分别通过阻断Tbx2和Sirt4的翻译来发挥其抑制心肌细胞肥大作用[13-14]。我们的实验证实miR-92b-3p通过靶向心肌细胞增强子2D(Mef2d)抑制心肌细胞肥大的发生[15]。我们前期miRNA表达谱芯片检测发现miR-199a-3p在血管紧张素 II(Ang-II)诱导肥厚的小鼠心肌中表达显著增强,但其对心肌肥厚的作用机制尚不明确。视网膜母细胞瘤转录辅阻遏子 1(retinoblastoma transcriptional corepressor 1,Rb-1)作为肿瘤抑制蛋白,通过抑制细胞周期进程,阻止细胞过度生长直至细胞准备分裂[16]。我们已证实cyclin/Rb-1信号通路激活介导了miR-16表达下调的促心肌细胞肥大作用[17]。本文利用乳小鼠心肌细胞,证实Rb-1是miR-199a-3p作用靶基因,并介导了miR-199a-3p的促小鼠心肌细胞肥大作用。
1 材料与方法
1.1 材料
限制性内切酶XhoI、EcoRI、转染试剂Lipofectamine 2000、Trizol、逆转录试剂盒、4 × SDS loading buffer(Invitrogen,Carlsbad,CA);2 ×SYBR Green Mix、RNAase free water(TaKaRa,Japan);miR-199a-3p mimic、Rb-1 siRNA(广州锐博);BCA蛋白定量试剂盒(Thermo Scientific,USA);SDS-PAGE凝胶试剂盒(Thermo Scientific,USA);抗 GAPDH、ACTA1,anti-Rabbit,antimouse抗体(Protein Technology,UK);抗ANP抗体(Bioworld,USA);抗β-MHC 抗体(Sigma,USA);蛋白 Marker(invitrogen,USA);PVDF膜(Millipore,USA);ECL发光液(Millipore,USA);Ang-II(Sigma,USA);DMEM/F12细胞培养基(Gibco,USA);特级澳洲胎牛血清(Gibco,USA)。其他生化试剂均为进口分装或国产分析纯。
1.2 乳小鼠心肌细胞培养及处理
取新生1~3 d的SPF级别C57BL/6乳鼠心脏(20只乳鼠/次,可铺1板12孔板细胞),以2.5 g/L胰蛋白酶消化法原代分离细胞。乳鼠心肌细胞(neonatal mouse ventricular cardiomyocytes,NMVC)与成纤维细胞因贴壁速度不同而得以分离,收集上清液中心肌细胞,将其接种于提前用1%明胶包被过的12孔板中,加入含有100 g/L胎牛血清及100 U/mL青霉素和100 mg/mL链霉素的DMEM/F-12培养基,置于37℃、体积分数5%CO2培养箱中培养。培养24 h后,更换一次完全培养基至稳定培养48 h。分别用100 nmol/L negative control、miR-199a-3p mimic和Rb-1 siRNA处理NMVC,24 h后结束实验。
1.3 FITC-鬼笔环肽(FITC-phallodin)染色
将NMVC种在confocal皿中,稳定生长后弃去培养基,用PBS漂洗两次,加入500 μL的40 g/L的多聚甲醛溶液固定,再用2 mg/mL的甘氨酸溶液中和多聚甲醛,摇床孵育2次,每次5 min。将10 μg/mL的FITC标记的鬼笔环肽染料37℃孵育40 min,用PBS漂洗2次,再加Hoechst33342反应液,避光37℃孵育40 min。将confocal皿倒扣于另一滴有30 μL防荧光淬灭封片剂的载玻片上,在倒置荧光显微镜下观察F-actin被染色后显示出细胞轮廓。
1.4 实时定量PCR检测Rb-1和miR-199a-3p的表达
用Trizol试剂提取心肌标本和NMVC总RNA。取1.0 μg总RNA,加入5×的逆转录试剂4 μL(逆转录试剂盒),用oligo(dT)15和random primers逆转录出cDNA用于检测编码基因mRNA水平。取1.0 μg总RNA,用miR-199a-3p特异的RT引物逆转录出cDNA用于检测miR-199a-3p水平。分别用GAPDH和U6作为检测编码基因和miR-199a-3p表达水平的内参照。在vii A7 Quantitative PCR System(Applied Biosystems,Carlsbad,CA)进行PCR反应和结果分析。以2-ΔΔCt法计算Rb-1和miR-199a-3p的相对表达水平。本文所用PCR引物序列见表1。
1.5 蛋白质印记迹法检测蛋白表达
收集心肌标本和处理后的NMVC,加入RIPA蛋白裂解液,冰上裂解,于4℃10 000×g离心10 min,取上清测浓度,并定量分装,加入4×上样缓冲液,99℃加热10 min使蛋白变性后,置于-80℃保存备用。蛋白样品进行聚丙烯酰胺凝胶电泳后,经电转至聚偏二氟乙烯(PVDF)膜上,用5%脱脂奶粉常温封闭1h,根据蛋白分子质量大小位置裁开,分别用相应的Ⅰ抗anti-ANP(1∶1 000)、anti-β-MHC(1∶1 000)、anti-α-actin-1(1∶1 000)、anti-Rb-1(1∶1 000)、anti-E2f2(1∶1 000)、anti-GAPDH(1∶5 000)4℃孵育过夜。TBST洗膜后,置于对应的II抗(1∶5 000)4℃孵育2 h。ECL发光试剂盒显影,应用ImageJ软件进行图像分析。

表1 PCR引物序列Table 1 The sequences of the primers for RT-qPCR
1.6 双荧光素酶报告基因实验验证miR-199a-3p与Rb-1 3'UTR的结合作用
参照我们已报道方法[18],分别构建包含miR-199a-3p潜在结合序列的Rb-1 3′UTR重组质粒pGL3-Rb-1-627-634及包含结合序列突变的重组质粒pGL3-Rb-1-627-634-MUT。HEK293细胞(细胞密度约为1×105个/孔/12孔板)转染200 ng重组质粒,50 nmol/L miR-199a-3p mimic以及10 ng pRL-TK(表达海肾荧光素酶的内参照质粒)。转染后24 h,测定萤火虫荧光素酶(firefly luciferase,FL)及海肾荧光素酶(renilla luciferase,RL)强度,两种荧光强度比值(FL/RL)变化可反映miR-199a-3p与Rb-1 3′UTR结合的能力。
1.7 统计学分析
应用SPSS 25.0统计软件,数据均采用均数±标准误(mean±SEM)表示,两组间比较采用t检验;多组间比较先进行正态分布和方差齐性检验,方差齐性检验后采用单因素方差分析(one way ANOVA),并用Bonferroni校正的t检验进行组间两两比较。P<0.05为差异有统计学意义。
2 结果
2.1 微小RNA miR-199a-3p促进心肌细胞肥大
原代分离乳小鼠心肌细胞,分别进行10 nmol/L Ang-II处理24 h或转染miR-199a-3p mimic。RT-qPCR结果显示,10 nmol/L Ang-II处理的NMVC中miR-199a-3p表达显著增加且差异有统计学意义(P <0.01;图1A);相对对照组 miR-199a-3p在NMVC水平显著升高(P<0.001;图1B),肥大相关基因Nppa,Acta1,Myh7表达显著增加(P<0.01;图1C)。同样的,Western blot结果显示,乳小鼠心肌细胞过表达miR-199a-3p后,相对对照组,肥大相关蛋白ANP,α-actin-1,β-MHC表达水平明显升高(P<0.05;图1D)。

图1 过表达miR-199a-3p促进小鼠心肌细胞肥大Fig.1 Overexpression of miR-199a-3p enhances cardiomyocyte hypertrophy
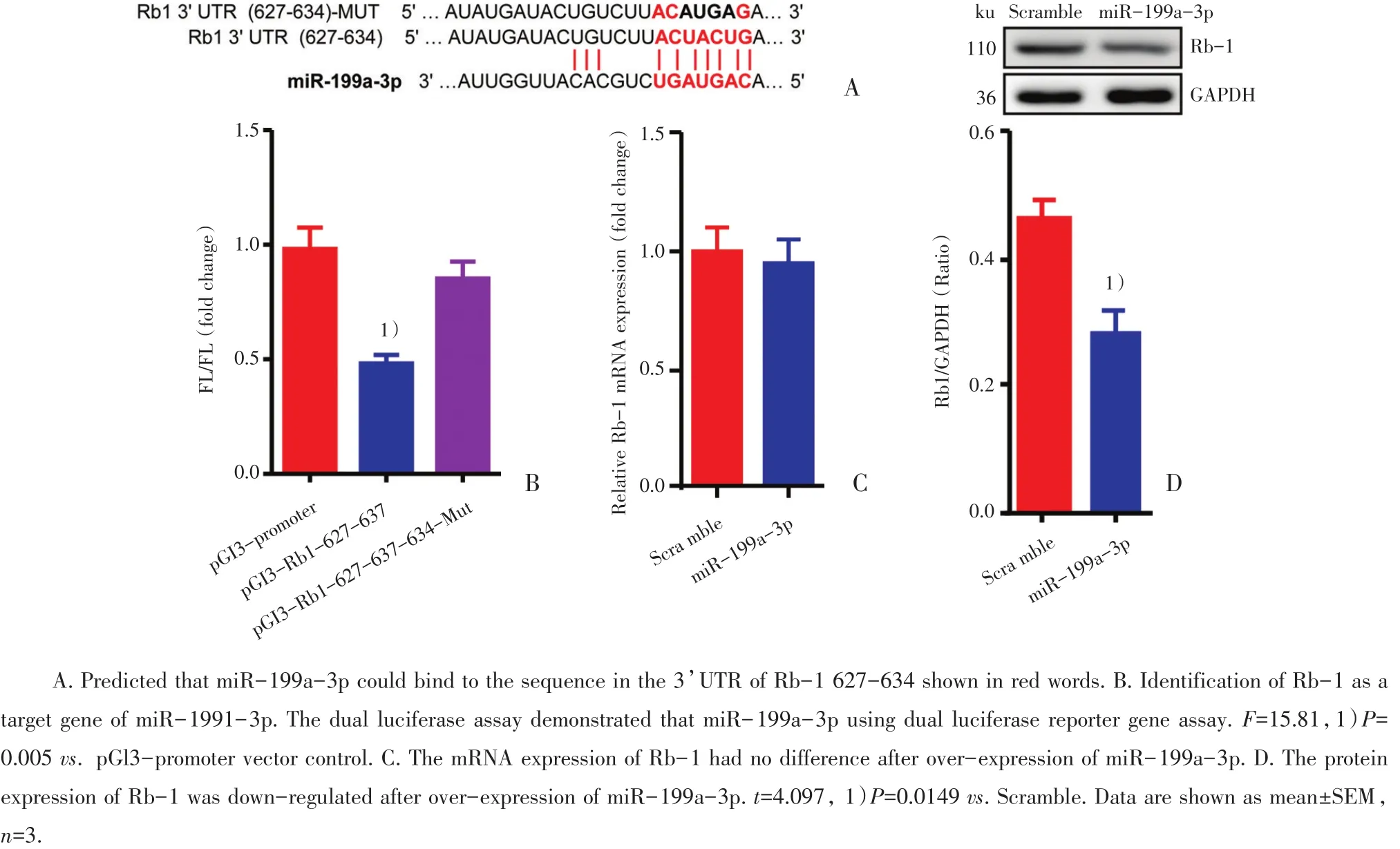
图2 MiR-199a-3p在转录后水平抑制Rb-1的表达Fig.2 MiR-199a-3p suppresses Rb-1 expression at the post-transcriptional level
2.2 双荧光素酶报告基因实验证实Rb-1与miR-199a-3p具有结合作用
基于Mirdb数据库(www.mirdb.org)以及TargetScan-Vert(www.targetscan.org)的序列分析提示,Rb-1 3’UTR的627-634碱基可能是miR-199a-3p潜在的结合位点(图2A)。双荧光素酶报告基因实验结果显示,与pGL3-promoter组相对比,重组质粒pGL3-Rb-1-627-634与miR-199a-3p mimic共转染组FL/RL值显著降低,差异具有统计学意义(P<0.01),而重组质粒pGL3-Rb-1-627-634-MUT与miR-199a-3p mimic共转染组FL/RL值无显著性差异(图2B)。RT-qPCR结果显示,增加乳小鼠心肌细胞中miR-199a-3p的水平后,相对对照组Rb-1的mRNA水平无明显差异(图2C)。Western blot结果显示,乳小鼠心肌细胞过表达miR-199a-3p后,Rb-1在蛋白水平表达明显下调,差异具有统计学意义(P<0.05;图2D)。这提示miR-199a-3p是在转录后水平抑制Rb-1的表达。
2.3 miR-199a-3p靶向Rb-1增加心肌细胞肥大
鬼笔环肽染色显示,乳小鼠心肌细胞过表达miR-199a-3p后,心肌细胞表面积明显增大;抑制Rb-1的表达,同样可以增加心肌细胞表面积,差异具有统计学意义(P<0.01;图3A)。在乳小鼠心肌细胞中过表达miR-199a-3p或者抑制Rb-1的表达,肥大相关蛋白ANP,α-actin-1,β-MHC表达水平明显增高,差异具有统计学意义(P<0.05;图3B)。乳小鼠心肌细胞过表达miR-199a-3p或者抑制Rb-1的表达,在细胞胞质内E2f2蛋白水平分别显著降低,差异具有统计学意义(P<0.05;图3C);而细胞核内的E2f2蛋白水平分别明显升高(P<0.01;图3D)。这表明miR-199a-3p通过抑制Rb-1的表达,促进了E2f2进入细胞核,从而发挥调控心肌细胞肥大相关基因表达的作用。
3 讨论
miR-199家族有三个成员,包括miR-199a1、miR-199a2和 miR-199b。miR-199a1和 miR-199a2分别位于19号和1号染色体上,具有相同的成熟序列[19]。miR-199a在压力超负荷的肥厚心脏中表达上调[20-22]。研究显示miR-199a抑制HIF-α表达,增加心肌细胞肥大[23]。MiR-199a也可以通过激活雷帕霉素(mTOR)复合物信号传导的哺乳动物靶点以细胞自主方式抑制心肌细胞自噬来促进心肌细胞肥大;而过表达Atg5或mTOR处理激活自噬增加了心肌细胞自噬可减轻由miR-199a过表达引起的心脏肥大[24]。
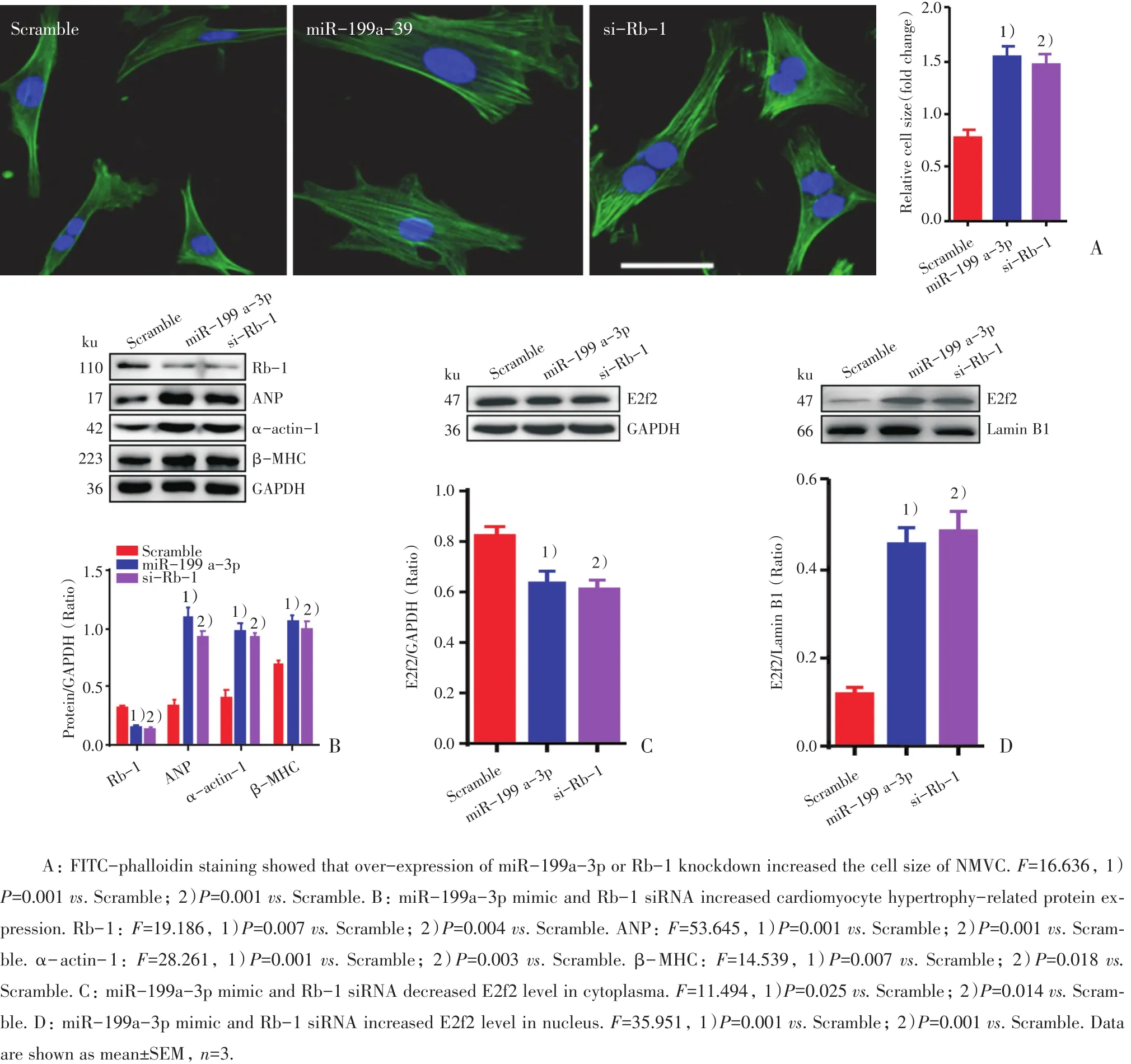
图3 E2f2入核增加介导miR-199a-3p促进心肌细胞肥大Fig.3 Increase of E2f2 transfer into nucleus mediates cardiomyocyte hypertrophy by miR-199a-3p
本文发现,在NMVC中过表达miR-199a-3p可以上调肥大相关基因表达,增加心肌细胞面积,与以往报道相符[23-24]。已有研究确定cyclin D2/pRb-1通路对于心肌肥大的调节有重要作用[25]。压力负荷诱发Rb-1基因敲除小鼠过度的心肌肥厚[26]。我们通过双荧光素酶报告基因实验证实了miR-199a-3p可与Rb-1 3’UTR具有结合作用,并且miR-199a-3p是在转录后水平抑制Rb-1的表达。抑制Rb-1表达能与miR-199a-3p一致性地促进心肌细胞面积增大和心肌细胞肥大相关的Anp、α-actin-1、β-MHC的表达增强。
已有研究表明,E2F2是一种广泛表达的转录因子,磷酸化的pRb导致E2F2的释放,从而促进细胞周期的进程[16]。E2F2作为Rb-1下游的转录因子参与对心肌细胞表型的调控作用[27-29]。在本文中,在NMVC过表达miR-199a-3p或者抑制Rb-1表达,E2f2在细胞内总蛋白水平表达一致性降低;但细胞核内的E2f2蛋白表达水平明显升高。这表明miR-199a-3p通过抑制Rb-1的表达,促进了E2f2进入细胞核,从而发挥促进心肌细胞肥大相关基因表达的作用。
综上,本文通过双荧光素酶报告基因实验、靶基因表达和相应功能性实验证实Rb-1是miR-199a-3p的作用靶基因;miR-199a-3p通过抑制Rb-1的表达,促进了E2f2进入细胞核,从而发挥促进心肌细胞肥大作用。在后续研究中,我们将在整体动物水平,进一步明确miR-199a-3p对Rb-1表达和心肌肥厚的调控作用。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
