青藏高原东缘高寒草甸土壤纤毛虫群落的季节变化特征
2019-02-26刘旻霞蒋晓轩李全弟南笑宁夏素娟
刘旻霞,蒋晓轩,李全弟,南笑宁,夏素娟
西北师范大学地理与环境科学学院,甘肃 兰州 730070
土壤纤毛虫的种类繁多、数量巨大,而且分布非常广泛,它们是土壤生态系统中很重要的组分(Giller,1996)。土壤纤毛虫与生态环境之间互相影响,对自然界的能量流通和元素转换过程中有显著作用(Bardgett et al.,2013)。而作为地下食物链和食物网的重要组成部分,土壤纤毛虫在凋落物的分解和土壤局部环境的改善中起到了很大作用(Liao et al.,2015)。土壤纤毛虫生存在地表环境系统中,因此它们在土壤生态系统中的个体数量和类群组成,与环境的变化密切相关(刘洁等,2017)。许多研究表明,土壤纤毛虫群落受气候(Stebaeva,2003;宋理洪等,2018)、植被(林英华等,2005)、土壤(王超等,2017)等生态因子的影响。随着温度、降水等生态因子的变化,不同地区土壤纤毛虫的群落组成与结构具有明显的地带性特征(尹文英,1992)。作为有效指示生物之一,土壤纤毛虫对不同季节及环境的变化极其敏感,故它们被广泛应用在其他生态系统(Petz et al.,1989;马正学等,2008)的评价中。
位于青藏高原东缘的甘南州,草场面积达2.723×106hm-2,高寒草甸是甘南地区分布较为广泛的生态系统类型,同时,也是甘南地区的重要草地资源,支撑着甘南地区的畜牧业发展,是黄河、长江的水源涵养区和补给区。近年来,由于全球气温升高、降雨量减少等自然因素和人口增加、草地过度放牧、旅游业的影响,全州草地生态环境日益恶化,天然草地大面积退化。甘南高寒草甸草场的退化和生境的改变等均会影响土壤纤毛虫个体、种群及群落结构的变化,反之,纤毛虫的这些改变也会影响局部生态环境及土壤理化性质(樊云龙等,2010;宁应之等,2017)。
由于高寒草甸地区四季环境因子变化明显,冬季漫长而寒冷,夏季多雨,降水集中,并且昼夜温差非常大,加之冬夏季草场放牧强度不均衡,导致土壤等环境因子变化巨大,这些都严重影响了土壤纤毛虫的生存环境。鉴于此,本文提出如下假设:(1)甘南高寒草甸土壤纤毛虫种类和数量随季节的不同而发生变化;(2)夏秋季土壤含水量增加,土温升高将增大土壤纤毛虫物种多样性。通过对以上问题的深入研究,总结高海拔气候变化敏感区土壤纤毛虫群落特征,以期为青藏高原高寒草甸牧区的土壤生物多样性保护和生态环境改善、退化生态系统的恢复提供基础数据。
1 研究地区与研究方法
1.1 研究区概况
甘南藏族自治州位于甘肃省西南部(34°55′N,102°53′E),地处青藏高原东缘,平均海拔3000 m。属高寒湿润类型,冷季长,暖季短,昼夜温差大,年均气温-0.5-3.5 ℃,极端最高气温28 ℃,极端最低气温-23 ℃。年均降水量558.8 mm,集中于7月、8月、9月。平均无霜期48 d,全年日照充足。地表径流深200-350 mm,年蒸发量1222 mm。土壤类型为亚高山草甸土,植被类型为亚高寒草甸,主要有优势植物三刺草(Aristida triseta)、矮嵩草(Kobresia humilis)、珠芽蓼(Polygonum viviparum)、金露梅(Potentilla fruticosa),伴生种有火绒草(Leontopodium alpinum)、鹅绒委陵菜(Potentilla anserina)等。
1.2 试验设计
在甘南合作市当周沟草原选取 3个样地(20 m×20 m,样地之间间距为200 m),分别于2015年4月(春季)、7月(夏季)、9月上旬(秋季)和12月初(冬季)在同海拔(3000 m左右)沿山头进行取样。同时测量土壤温度、照度及含水量,并调查植被状况。在每个样地中设置 9个样方(图1),用直径为5 cm的土钻沿对角线分别取0-5、5-15、15-25 cm土层土壤,装入自封袋,并贴好标签,4个季节共取得土样108份,带回实验室,待分析。
图1 实验样地分布图Fig.1 Plot distribution map
1.3 项目的测定与方法
1.3.1 土壤因子测定
采用烘干法测定土壤含水量;用地温计(EM50)测定土壤温度,以上实验均做3次重复。用型号为ZDS-10的照度计测量各样地光照度,测量时照度计和地面保持水平,测量高度距离地面50 cm。土壤有机碳采用重铬酸钾容量法测定;土壤全氮(TN)消煮后,采用硫酸-K2SO4∶CuSO4∶Se 催化法消煮,使用SmartChem 200化学分析仪进行测试。土壤全磷(TP)及速效磷(AP)采用钼锑抗法比色法测定(Olsen et al.,1954)含量;pH值采用电位法测定;速效氮(AN)用 KCl浸提取上清液,采用SmartChem 200化学分析仪进行测试。
1.3.2 样品培养与鉴定
采用“非淹没培养皿法”(Foissner et al.,2002)鉴测土壤纤毛虫。把土壤浸出液加入有土样的培养皿中,将土壤充分润湿,但不淹没,在 25 ℃恒温下培养土壤纤毛虫,培养开始后每隔3 d置于光学显微镜下鉴定土壤纤毛虫物种,直到没有新物种出现为止。采用活体观察法和染色制片法鉴定土壤纤毛虫物种。
1.4 数据统计与分析
土壤纤毛虫的多样性特征分析采用以下各指数计算公式(Whittaker,1972):
(1)多样性指数(Shannon-Wiener)公式:

(2)均匀度指数(Pieluo)公式:

(3)优势度指数(Simpson)公式:

式中,H为物种的多样性指数;Pi为第i类群的个体数比,Pi=Ni/N,Ni为第 i类群的个体数,N为总个体数;S为所有类群数。
不同季节土壤纤毛虫相似性用 Jaccard相似性指数公式计算:

式中,当 0.50≤q<0.7时,表示中等相似;当0.25≤q<0.50 时,表示中等不相似;0<q<0.25 时,表示极不相似(宁应之等,1998)。
数据的统计分析运用Excel 2013软件进行,运用SPSS 18.0软件进行One-way ANOVA(单因素方差)分析,运用Canoco for windows 4.5进行RDA(Redundancy analysis)排序分析。运用SigmPlot 12.0软件作图。
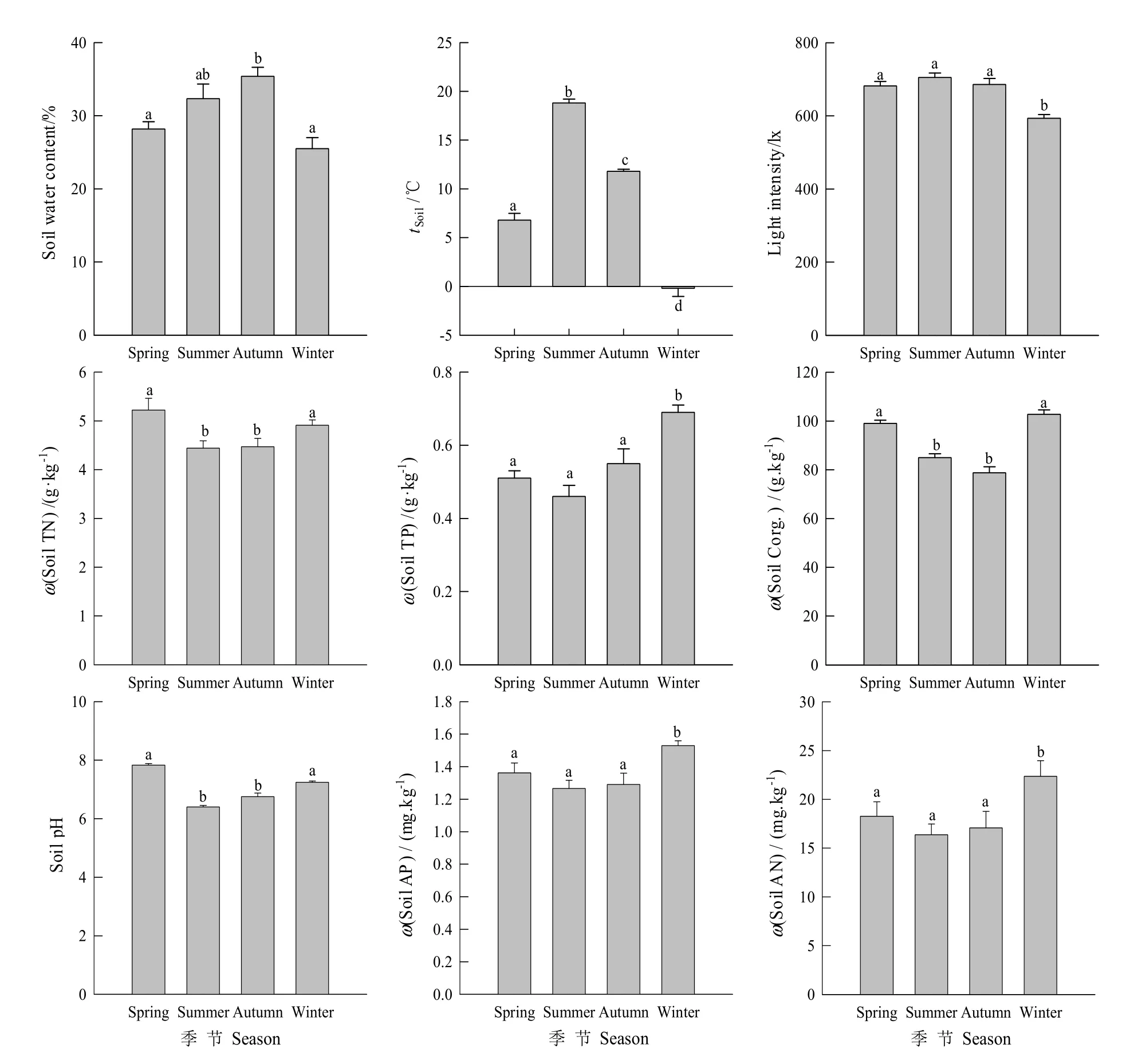
图2 土壤环境因子的四季变化Fig.2 Changes of soil environmental factors in the four seasons
2 结果与分析
2.1 环境因子的四季变化特征
图2表明,研究区土壤含水量以秋季最高,其次是夏季,春冬季较小,不同季节之间存在显著差异(P<0.05);土壤温度表现为夏季>秋季>春季>冬季,四季差异显著(P<0.05);光照度除冬季较低外,其他3个季节差异不显著;土壤全氮(TN)、全磷(TP)、有机碳(SOC)、速效氮(AN)及速效磷(AP)含量在不同季节间差异显著(P<0.05),春冬季土壤有机碳显著高于其他两个季节,这可能是由于冬春季土壤凋落物分解所致;而土壤pH春季最高,冬季次之,夏秋季相对较低,这与四季降水的不均衡有关。
2.2 土壤纤毛虫的群落组成
图3显示:在研究区春季土壤中共鉴出97种纤毛虫,分属于9纲17目32科49属,其中旋毛纲(Spirotrichea)和裂口纲(Litostomatea)为优势类群,分别有16属26种和7属20种。夏季土壤纤毛虫有141种,分属于9纲17目32科55属,其中旋毛纲和裂口纲为优势类群,分别有18属40种和10属28种。秋季土壤中共有105种纤毛虫,分属于9纲17目32科48属,旋毛纲和裂口纲为优势类群,分别有16属26种和9属22种。冬季共有纤毛虫78种,分属于9纲17目32科42属,旋毛纲和裂口纲为优势类群,分别有13属21种和8属15种。土壤纤毛虫的丰度四季变化明显,秋季的丰度高于夏季和冬季,春季稍低一些。
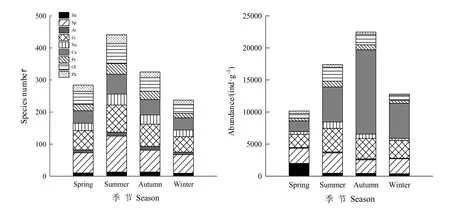
图3 土壤纤毛虫群落的四季变化Fig.3 Changes of soil ciliates community in four seasons

图4 不同季节土壤纤毛虫多样性指数Fig.4 Diversity index of soil ciliates in different seasons
2.3 土壤纤毛虫群落多样性与相似性
Margalef丰富度指数(4.64)和Pieluo均匀度指数(0.98)均是春季相对较高,其他季节相对较低。Shannon-Wiener多样性指数(10.56)、Simpson优势度指数(0.26)、物种数(91)和丰度(5332)均是夏季最高,其他季节相对较低,其中丰度以冬季最低(256)(图4)。
土壤纤毛虫群落的相似性指数在 0.546-0.738之间(表1),其中夏秋季相似性指数最高(0.738),春夏季次之(0.688),为中等相似,冬夏季相似性指数较低(0.546)。

表1 土壤纤毛虫群落相似性分析Table1 Similarity analysis of soil ciliates community

图5 不同季节土壤纤毛虫种类数量的垂直分布Fig.5 Species number and abundance distribution of soil ciliates in different seasons
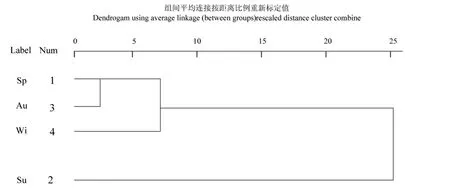
图6 土壤纤毛虫种群在四季分布的聚类分析Fig.6 Cluster analysis of the distribution of soil ciliates in four seasons
2.4 土壤纤毛虫垂直分布动态
随土层加深,4个季节纤毛虫种类数和丰度都呈逐渐减少的趋势,表明土壤纤毛虫主要分布在表层土壤,中层和深层分布较少。夏季和秋季的物种数和丰度较冬春季高,4个季节的物种数和个体数差异显著(P<0.05)。不同季节间,0-5 cm的物种数以夏季最高(63.4),个体数则以秋季最高(3214),春季的类群数(43.4)和个体数(794)相对较低。5-15 cm 的物种数(48.2)和个体数(1808)均以夏季为最高,秋季次之,春(37.4和556)冬(28.2和590)季较低。与表中层相比,10-15 cm土层的物种数和个体数波动幅度不大(图5)。高寒草甸土壤纤毛虫物种数和个体数的垂直分布均有一定的表聚性,但表聚性程度因季节而异。
2.5 不同季节土壤纤毛虫聚类分析
根据不同季节土壤纤毛虫的类群数进行分析,结果如图6所示,春季和秋季最先聚为一类,随后冬季和春季聚为一类后与夏季聚为一类,表明春秋季的土壤纤毛虫分布最为相似,夏季差异最大,相似度表现出随季节变化、环境变化及纤毛虫种类数量的变化而变化的特征(图6)。
2.6 土壤纤毛虫与不同季节环境因子关系
采用RDA(冗余分析)方法进行相关分析,图7中箭头(实线)所在的位置代表排序轴和环境因子之间的相关性(正负),其箭头和排序轴的夹角大小表示环境因子的相关性大小。结果显示,大部分的纤毛虫物种分布在夏秋两季,尤其以夏季居多。纤毛虫与土壤含水量、光照度、土壤温度呈显著正相关,与其他环境因子呈负相关关系,且正相关性大小表现为:土壤温度>光照度>土壤含水量。由不同科土壤纤毛虫冗余分析的第一、二排序轴和环境因子的相关系数可知,第一轴排序轴解释信息量为83.6%,第二排序轴为8.9%。第一、二排序轴累积可以解释信息量达92.5%,可以说明前两个排序轴反映了环境因子与纤毛虫结构的大部分信息(表2)。
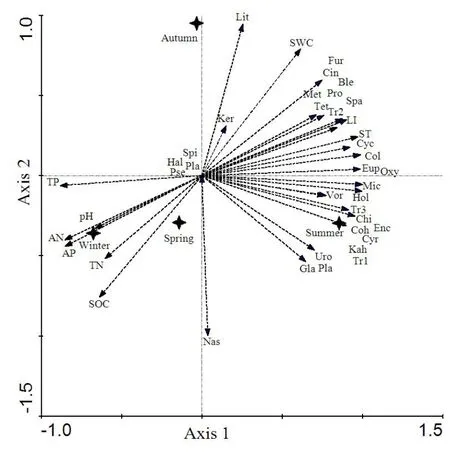
图7 不同季节土壤纤毛虫物种与环境因子的冗余分析Fig.7 RDA of soil ciliate species and environmental factors in different seasons
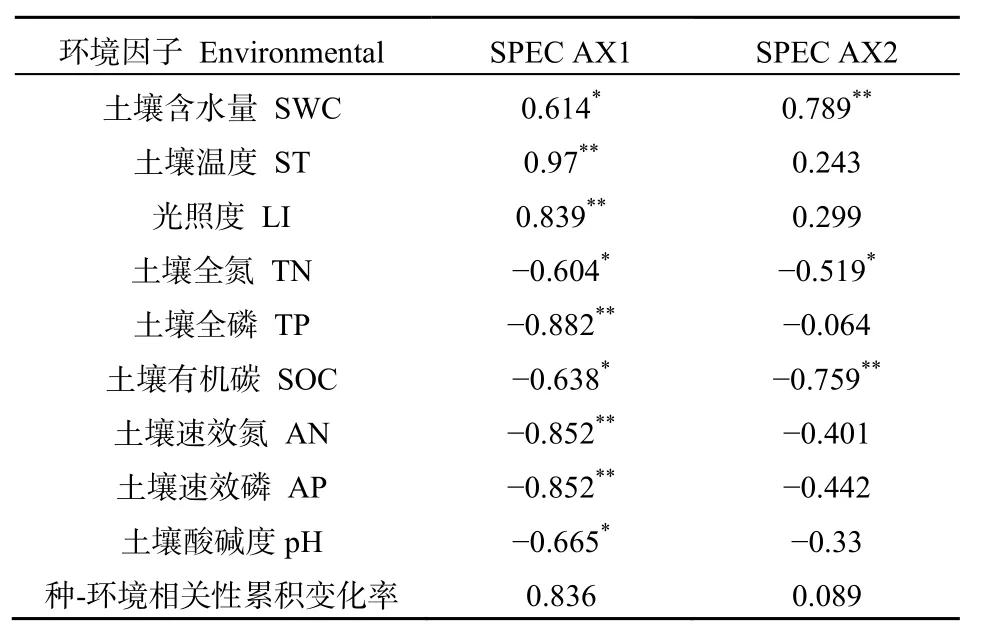
表2 排序轴与环境因子的冗余分析Table2 RDA analysis of ordination axis and environmental factors
3 讨论
不同季节土壤温度、光照度、土壤含水量及土壤养分含量等外界条件的差异会影响土壤纤毛虫的群落结构、多样性等的变化。本研究中,土壤养分、含水量、光照度、土温及pH等在不同季节间有显著差异(图 2),其中,土壤温度的四季差异最为显著,光照度冬季偏低,土壤含水量夏秋季高于其他两季,土壤养分含量在植物生长的旺季相对较低,主要原因是甘南地区夏秋季降水较多,而过多的降水会造成土壤养分尤其是表层养分流失严重。除此之外,夏秋季植物生长较为旺盛,为了满足其自身生长发育对养分的需求,植物根系同样需要吸纳大量的养分,从而导致土壤养分损耗较多,养分含量呈下降趋势,这与其他相关研究(薛敬意等,2003)结果一致。土壤有机碳含量在春冬季较高,这可能与植物凋落物回归有关,生长旺盛期的土壤pH值低于其他季节,这与江远清等(江远清等,2007)的发现基本吻合,说明土壤pH值的变化是一个相对复杂的过程,土壤中 H+浓度容易受土壤温湿度及其他阳离子等的影响(Baldrian et al.,2013)。
甘南高寒草甸土壤纤毛虫群落具有显著的季节变化特征,印证了本文提出的第一个假设。4个季节中物种数和丰度在夏秋季最多,而旋毛纲和裂口纲为优势类群(图3)。土壤动物季节动态分布受温度、降水和植被等的影响而出现波动(Morónríos et al.,2010),不同地区土壤动物多样性的季节动态不同。有研究表明,土壤动物的类群数和密度随土温的升高而减小,然而在长白山地区随土壤温度的升高,土壤动物类群数和密度呈增大趋势(李红月,2015);另有研究发现,土壤动物的个体数、类群数及多样性在雨季最低(杨效东等,2003)。在寒温带和中温带地区土壤动物密度在 7-9月达到最大;在夏秋季节(雨热同期)的土壤动物群落多样性和数量都较高,相反冬春季就较少(殷秀琴等,2010;李玉杰等,2018)。多样性指数(Shannon Wiener)、均匀度指数(Pielou)在春季最高;优势度指数(Simpson)、丰富度指数(Margalef)、物种数和丰度在夏季最高(图4),这可能与四季的环境因子变化有关。春季土温,降水及养分都比较适中,适合大部分纤毛虫的生长,所以土壤纤毛虫的多样性较高;而夏秋季降水较多,土壤淋溶以及地上植被快速增长消耗碳氮磷等养分,除此之外降水过多也会改变土壤中溶液的渗透压、破坏土壤团粒结构,使土壤纤毛虫及微生物等的生命活动减弱(戴允泽等,2018)。细菌、轮虫和动植鞭毛虫等物种数的减少无疑会影响到一些对食物有选择性的纤毛虫的食物来源(齐莎等,2010)。这样就会导致部分纤毛虫过度生长而竞争排除掉其他物种,使少数纤毛虫成为优势度,从而使优势种和丰度等相应增加,这与本文提出的第二个假设不一致。土壤纤毛虫群落相似性(表 1)显示,4个季节之间,夏秋季相似性指数最高(0.738),春夏季次之(0.688),均为中等相似,冬夏季相似性指数较低(0.546)。这可能与各个季节的环境因子影响有关。群落结构的复杂性和各种群数量受到环境因子和生物群落适应能力的双重影响。
高寒草甸生态系统土壤纤毛虫类群数和个体数的垂直分布均有一定的表聚性,但不同季节表聚性程度不同(图5)。已有研究表明,土壤动物的垂直分布受土壤理化性质和营养状况的影响(肖能文等,2009)。草原生态系统表层土壤是植物根系的主要分布区,土壤相对疏松,通气性好,营养丰富因而土壤动物相对较多。植被结构和土壤理化性质是决定土壤动物垂直分布的相对稳定性因子。
RDA进一步分析结果表明(图7),四季环境因子均对土壤纤毛虫的分布产生了影响,其中土壤温度的影响最大。这与其他研究结果一致,即土壤温度和大气降水导致的土壤温湿度变化是引起原生动物种群产生季节差异的主要原因(王薪琪等,2017)。土壤纤毛虫生存最适宜的温度是地表温度在14.3-24.3 ℃范围(宁应之等,2017)。处于甘肃省南部的甘南州草原冬季漫长而寒冷,进入夏秋季后雨量增加,气候变暖,土壤湿度增加(图2),土壤动物繁殖能力增强。本研究中,与其他 3个季节相比,土壤温度和含水量在夏季最高,这与宁应之等(2017)的研究结果一致。另外,夏季土壤温度和湿度的变化还可以影响植物的生理和群落结构,从而影响土壤原生动物的资源可得性和微生境。
4 结论
(1)甘南高寒草甸随着季节的变化,降水、温度及土壤养分等不同,土壤纤毛虫群落多样性表现出显著不同。物种丰富度、多样性、物种数等总体来说在夏秋季较高,分别为:5332,10.56及91;冬季较低,分别为:256,5.9及47.4,体现了最优良的生境和最大的物种多样性指数。
(2)土壤温度、全磷含量、光照度和含水量等均影响了甘南高寒草甸土壤纤毛虫群落的分布,其中土壤温度是关键限制因子。
(3)土壤纤毛虫物种数和丰度垂直分布明显,均为0-5 cm>5-15 cm>15-25 cm,这体现了纤毛虫分布的表聚性。
