小麦抗逆相关转录因子DREB密码子偏好性特征分析
2019-02-25雷梦林冯瑞云郝雅萍王慧杰杨生权
雷梦林,冯瑞云,郝雅萍,刘 霞,王慧杰,杨生权
(1.山西省农业科学院农作物品种资源研究所/农业部黄土高原作物基因资源与种质创制重点实验室/杂粮种质资源发掘与遗传改良山西省重点实验室,山西太原 030001; 2.山西省农业科学院作物科学研究所/作物遗传与分子改良山西省重点实验室,山西太原 030031; 3.山西省生物研究所,山西太原 030006)
干旱、高盐、低温等是影响作物产量提高的主要非生物胁迫因素[1]。DREB转录因子是一个应答非生物逆境的重要结合蛋白,能特异结合DRE/CRT顺式元件,并调控下游一系列抗逆基因的转录表达,从而增强植物的耐逆性。目前,已从拟南芥[2]、水稻[3]、苹果[4]、山葡萄[5]、大豆[6]、玉米[7]和小麦[8]等植物中分离到DREB类基因,但该基因最佳外源表达系统的选择和表达水平的预测等问题有待解决,分析密码子的使用偏好性有助于指导其异源表达和遗传转化等后续研究的开展。
密码子是联系基因核苷酸序列与蛋白质氨基酸序列的途径,在生物体遗传信息的传递过程中发挥着关键性作用。密码子偏性[9]是指某一特定基因、物种对某种或多种同义密码子的使用偏好,普遍存在于生物体中,并与特定基因的碱基组成、表达水平等因素关联。进行密码子的偏性分析有助于预测特定基因的功能及其表达调控机制和分子进化水平。近年来,有关密码子偏性在水稻[10]、拟南芥[11]、葡萄[12]等植物中均有研究,还未见小麦DREB基因密码子使用偏性的相关报道。本研究拟通过CodonW和EMBOSS在线程序,分析小麦DREB密码子偏性,并与其他13种植物的DREB密码子偏性进行比较,以明确物种间的亲缘关系与密码子使用频率之间的关系;最后与大肠杆菌、酵母、拟南芥、烟草和番茄基因组密码子偏性进行比较,为DREB基因在转基因研究中选择最佳受体植物提供参考,也为深入开展DREB的蛋白质结构、功能和转基因功能验证提供基础数据。
1 材料与方法
1.1 序列数据来源
小麦DREB mRNA克隆于中国春,序列全长1 088 bp,其中ORF长度837 bp,编码278个氨基酸,GenBank(https://www.ncbi.nlm.nih.gov/)登录号为AF303376.1。
玉米、拟南芥、滨藜、燕麦、榛子、大豆、亚洲棉、黄花苜蓿、水稻、碱蓬、刚毛柽柳、橡胶草和中间偃麦草等13种植物的DREB序列来源于GenBank(表1)。
大肠杆菌和酵母基因组的密码子偏性数据来源于Codon Usage Database(http://www. kazusa. or. jp/codon)。
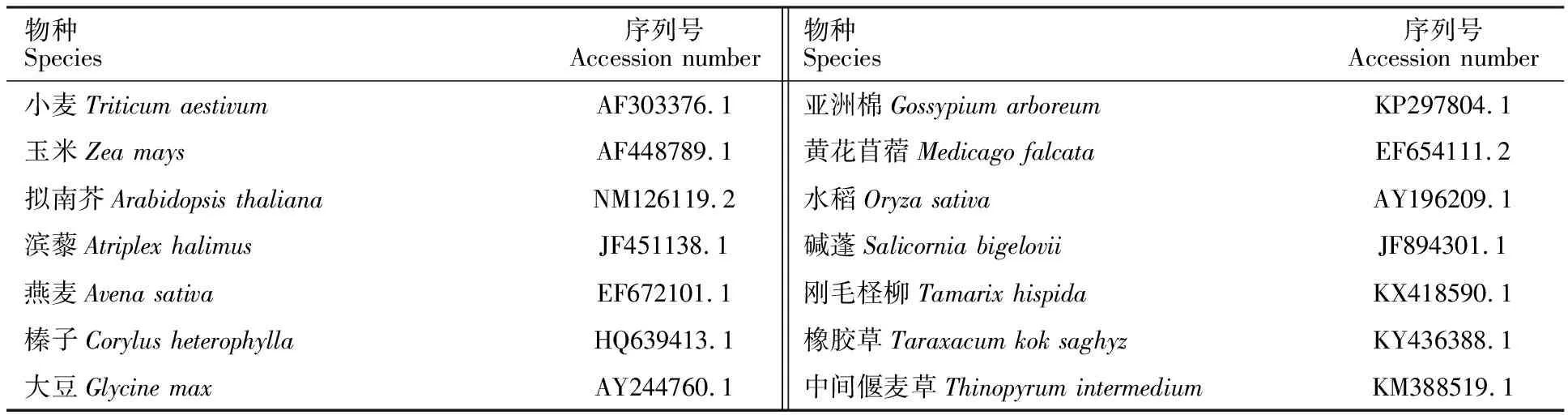
表1 DREB基因的编码区序列来源Table 1 Sources of coding sequence of DREB genes
1.2 密码子偏性分析
密码子的使用频率采用CodonW软件和EMBOSS在线程序(http://vmbioinfo. toulouse. inra. fr/emboss) 中的CHIPS(Codon heterozygosity in a protein coding sequence)、CUSP(create a codon usage table) 分析。
1.3 同义密码子使用偏性分析
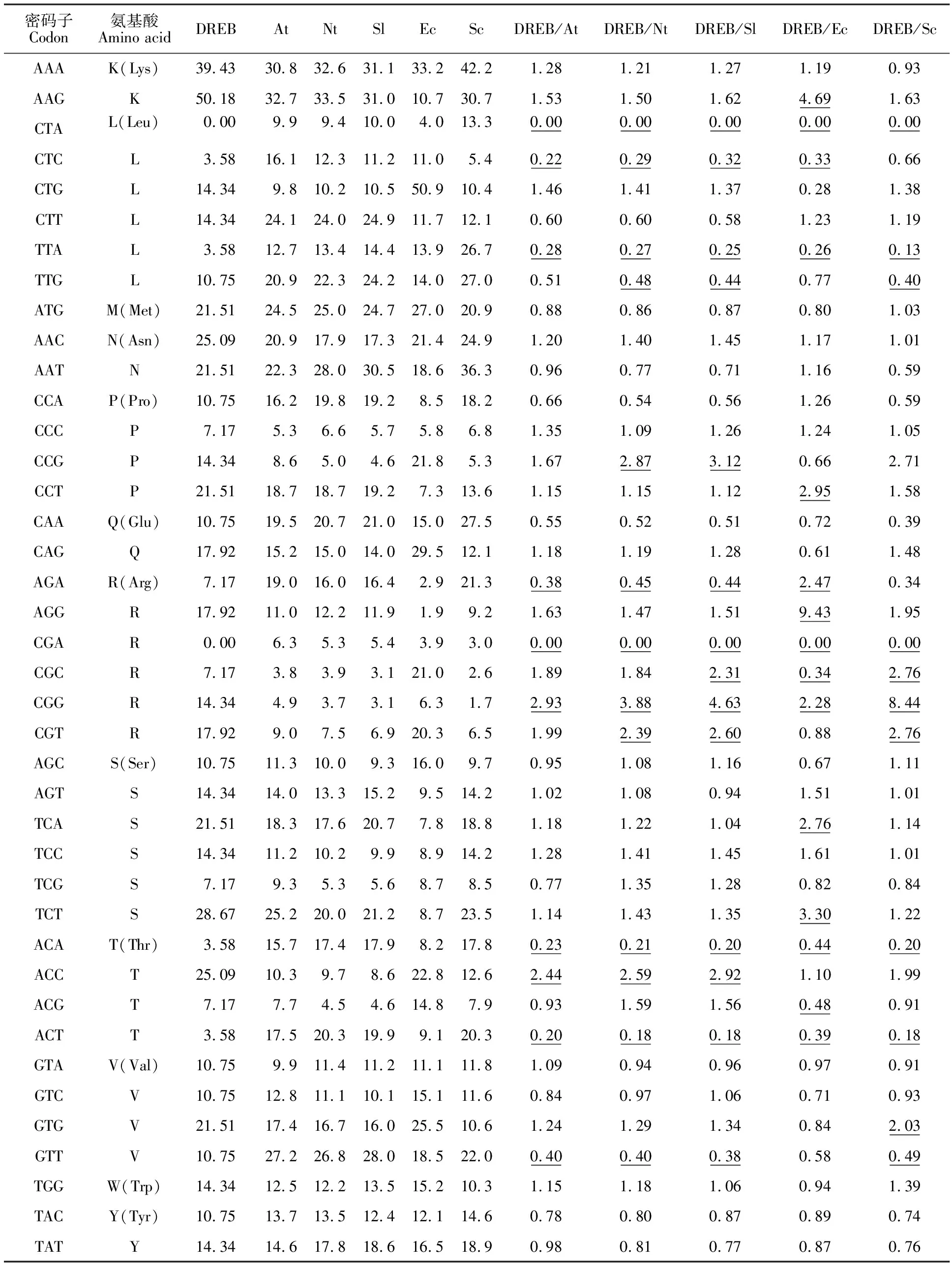
密码子使用的偏性程度用同义密码子的相对使用度(relative synonymous codon usage,RSCU)评价,RSCU值=某一密码子所使用的频率/其在无偏使用时预期频率,若RSCU值=1,则表明该密码子的使用没有偏好;RSCU值>l,表明该密码子使用频率相对较高;RSCU值 基因的碱基组成偏好性通过GC3s来评价,GC3s=密码子的第3位碱基中GC的含量/第3位碱基总量[14]。 密码子使用频率是衡量物种间密码子使用偏性差异的指标之一,为各密码子占编码该氨基酸的密码子的比例,比值在0.5~2.0之间表示二者的密码子偏好性接近,比值≤0.5或比值≥2.0,表示二者的密码子偏性差异较大[15]。 基因整体密码子偏性用有效密码子数(effective number of codons,ENc)评价,其值在20~61之间,越靠近20偏性越强,而值越大,其基因对密码子的选择偏性相对较弱,且稀有密码子增多[16]。 编码区同义密码子与密码子最佳使用相符合的程度通过密码子适应指数(codon adaptation index,CAI)来反映,取值范围在0~1之间。 将14个DREB转录因子的密码子使用偏性利用SPSS 18.0软件进行聚类分析。对基因密码子使用概率分析时,将每一个基因定义为一个对象。采用RSCU的欧氏平方距离表示基因间的距离,采用离差平方和法[17]表示类与类间的距离。DREB基因CDS编码区序列系统发育树是利用MEGA 5.1软件并结合组间邻接法(neighbour joining)来构建。 2.1.1 小麦DREB的 ENc、GC、GC3s和CAI分析 运用CodonW和CHIPS在线程序,计算得出小麦DREB的ENc值、GC含量和GC3s值分别为54.00、0.518和0.481,其中,ENc值大于35,偏向于61,表明在小麦中该基因对密码子选择偏性比较弱,且表达水平偏低(表2)。DREB编码区GC含量为51.8%,基因编码区序列中AT(48.2%)与GC含量(51.8%)含量基本相当,略微偏好于使用以GC结尾的密码子。小麦DREB的GC3s为0.481,表明了小麦DREB密码子第三位碱基偏好使用以AT结尾。小麦DREB的CAI值为0.262,远小于1,进一步表明小麦DREB对密码子的偏好性偏弱,可能在不同组织中的表达水平普遍偏低。 表2 不同物种DREB的ENc、GC、GC3s和CAI分析Table 2 Analysis of ENc,GC,GC3s and CAI for DREB in different species 2.1.2 DREB同义密码子相对使用度 运用CodonW和EMBOSS在线程序分别计算各基因的RSCU值和密码子使用频率,结果表明,小麦DREB基因密码子中,RSCU值>l的有25个密码子,RSCU值>l.5的有14个密码子,其中,密码子ACC的RSCU值达到2.550,偏好性最强;而TGG的RSCU值为1,表明该密码子不具有使用偏好性,可以随机被选用。I(Ile)、V(Val)、K(Lys)、Y(Tyr)、Q(Gln)、N(Asn)、E(Glu)和A(Ala)8种氨基酸的密码子偏好性相同;F(Phe)、H(His) 和D(Asp) 3种氨基酸的同义密码子偏性完全相反(表3)。 2.2.1 不同物种ENc、RSCU及GC含量分析 表3 小麦DREB的密码子偏性分析Table 3 Analysis of codon bias of DREB in Wheat 14个物种DREB的ENc平均值为52.770,其中,水稻的ENc为34.160,小于35;其余13个物种ENc值均大于35,说明这些物种的密码子偏好性及基因表达水平虽有一定差异性,但密码子选择偏好性和基因表达水平普遍较低,与CAI指数验证结果相同。滨藜的ENc值为61,表明其密码子碱基选择不具有偏好性。14个物种GC3s值介于0.304~0.853,均值为0.518;GC含量值介于0.414~0.724,均值为0.527。不同物种DREB对GC和GC结尾的密码子的偏好性不同,其中,4个双子叶植物(大豆、黄花苜蓿、碱蓬、橡胶草)GC含量小于0.5,偏向于AT结尾的密码子,而5种单子叶植物的DREB和剩余5种双子叶植物均对GC有一定的偏好性,其中,单子叶植物玉米对GC的偏性最强。利用RSCU值分析14个物种DREB密码子使用偏好性,结果表明,RSCU值>1的密码子数目最多的是榛子,为30个;最少的是刚毛柽柳,为20个。RSCU值>1.5的密码子数目最多的是水稻,为21个;最少的是刚毛怪柳,为6个(表2)。 2.2.2 密码子选用偏好性和CDS序列的系统聚类 通过对14个不同物种DREB基因的密码子使用偏性进行聚类分析。结果(图1)表明,14个物种可聚为2个大类,单子叶植物玉米和水稻属于Ⅰ类,剩余的12种属于Ⅱ类。在Ⅱ类中,单子叶植物小麦、中间堰麦草和燕麦单独归为一小类。单子叶植物玉米和水稻与单子叶植物小麦、中间堰麦草和燕麦分别属于禾本科的不同类群,说明亲缘关系较近的物种密码子使用频率也相似。而同属于藜科的碱蓬和滨藜没有聚为一小类,可见物种之间DREB的密码子使用偏性与物种之间进化并不一定有必然的联系。双子叶植物同属于Ⅱ类,说明双子叶植物DREB基因的密码子偏好性相对一致,而单子叶植物内部在长期进化过程中逐渐朝不同的方向进化,存在更为丰富的密码子偏好性。 14个不同物种DREB基因CDS序列构建系统发育树的分析结果(图2)表明,14个物种基本按照单子叶植物和双子叶植物进行分类,禾本科的单子叶植物小麦、中间堰麦草和燕麦与禾本科的单子叶植物玉米和水稻各归为一个小类。与密码子偏性的聚类分析结果相比,藜科的碱蓬和滨藜从不同类归到同一类中,并且进化距离较小。分属于不同科的双子叶植物归类可能是由于研究样本数量较少,导致不同科归于同一个分类。 通过将小麦DREB与拟南芥、烟草、番茄、大肠杆菌、酵母菌基因组中不同密码子出现的频率进行比较分析发现(表4),小麦DREB与大肠杆菌、酵母菌分别有26和22个密码子存在较大的选用偏好性差异,说明酵母菌真核异源表达系统更适用于小麦DREB异源表达试验。分析比较小麦DREB与遗传转化模式植物拟南芥、烟草和番茄的密码子偏好性发现,与拟南芥、烟草和番茄基因组密码子选用频率差异较大的密码子个数分别为16、22和24,表明在小麦DREB基因遗传转化功能验证中,拟南芥更适用于异源受体。 图1 基于DREB密码子使用偏性的聚类分析 欧氏距离 Euclidean distance 图2基于DREB密码子CDS序列的系统聚类 Fig.2ClusteranalysisdendrogramofCDSsequencesofDREB 表4 小麦DREB基因与部分模式生物基因组密码子使用偏性比较Table 4 Comparison of codon usage preference between DREB in Triticum aestivum and other representative organisms (续表4Continuedtable4) 密码子Codon氨基酸Amino acidDREBAtNtSlEcScDREB/AtDREB/NtDREB/SlDREB/EcDREB/ScAAAK(Lys)39.4330.832.631.133.242.21.28 1.21 1.27 1.19 0.93 AAGK50.1832.733.531.010.730.71.53 1.50 1.62 4.691.63 CTAL(Leu)0.009.99.410.04.013.3 0.00 0.00 0.00 0.00 0.00CTCL3.5816.112.311.211.05.4 0.22 0.29 0.32 0.33 0.66 CTGL14.349.810.210.550.910.41.46 1.41 1.37 0.28 1.38 CTTL14.3424.124.024.911.712.10.60 0.60 0.58 1.23 1.19 TTAL3.5812.713.414.413.926.7 0.28 0.27 0.25 0.26 0.13 TTGL10.7520.922.324.214.027.00.51 0.48 0.44 0.77 0.40 ATGM(Met)21.5124.525.024.727.020.90.88 0.86 0.87 0.80 1.03 AACN(Asn)25.0920.917.917.321.424.91.20 1.40 1.45 1.17 1.01 AATN21.5122.328.030.518.636.30.96 0.77 0.71 1.16 0.59 CCAP(Pro)10.7516.219.819.28.518.20.66 0.54 0.56 1.26 0.59 CCCP7.175.36.65.75.86.81.35 1.09 1.26 1.24 1.05 CCGP14.348.65.04.621.85.31.67 2.87 3.12 0.66 2.71 CCTP21.5118.718.719.27.313.61.15 1.15 1.12 2.951.58 CAAQ(Glu)10.7519.520.721.015.027.50.55 0.52 0.51 0.72 0.39 CAGQ17.9215.215.014.029.512.11.18 1.19 1.28 0.61 1.48 AGAR(Arg)7.1719.016.016.42.921.3 0.38 0.45 0.44 2.470.34 AGGR17.9211.012.211.91.99.21.63 1.47 1.51 9.43 1.95 CGAR0.006.35.35.43.93.0 0.00 0.00 0.00 0.00 0.00 CGCR7.173.83.93.121.02.61.89 1.84 2.310.34 2.76CGGR14.344.93.73.16.31.72.933.884.632.28 8.44CGTR17.929.07.56.920.36.51.99 2.392.600.88 2.76AGCS(Ser)10.7511.310.09.316.09.70.95 1.08 1.16 0.67 1.11 AGTS14.3414.013.315.29.514.21.02 1.08 0.94 1.51 1.01 TCAS21.5118.317.620.77.818.81.18 1.22 1.04 2.76 1.14 TCCS14.3411.210.29.98.914.21.28 1.41 1.45 1.61 1.01 TCGS7.179.35.35.68.78.50.77 1.35 1.28 0.82 0.84 TCTS28.6725.220.021.28.723.51.14 1.43 1.35 3.301.22 ACAT(Thr)3.5815.717.417.98.217.80.230.210.20 0.44 0.20ACCT25.0910.39.78.622.812.62.442.592.92 1.10 1.99 ACGT7.177.74.54.614.87.90.93 1.59 1.56 0.480.91 ACTT3.5817.520.319.99.120.30.200.180.180.390.18GTAV(Val)10.759.911.411.211.111.81.09 0.94 0.96 0.97 0.91 GTCV10.7512.811.110.115.111.60.84 0.97 1.06 0.71 0.93 GTGV21.5117.416.716.025.510.61.24 1.29 1.34 0.84 2.03GTTV10.7527.226.828.018.522.00.400.400.380.58 0.49 TGGW(Trp)14.3412.512.213.515.210.31.15 1.18 1.06 0.94 1.39 TACY(Tyr)10.7513.713.512.412.114.60.78 0.80 0.87 0.89 0.74 TATY14.3414.617.818.616.518.90.98 0.81 0.77 0.87 0.76 At、Nt、Sl、Ec、Sc分别表示拟南芥、烟草、番茄、大肠杆菌、酵母菌;下划线表示两个物种密码子比较具有明显偏差(≤0.5,≥2.0)的分值。 At,Nt,Sl,Ec,and Sc representArabidopsisthaliana,Nicotianasylvestris,Solanumlycopersicum,EscherichiacoliandSaccharomycescerevisiae. The underlines mean there are obvious differences between the codons of two species(≤0.5,≥2.0). 生物在长期的进化过程中,会形成一套使核苷酸和氨基酸之间保持遗传信息传递的特定密码子用法,由于基因的突变和选择等因素的影响[18-21],自然选择通常使生物体偏向于使用最优密码子,突变会导致部分非最优密码子存在,经过长期进化,同一物种或不同物种的不同基因对密码子的使用表现为不同的偏好性。 本研究通过对小麦DREB的密码子偏性分析,发现该基因碱基选择时偏好GC,但其密码子第3位碱基偏好AT结尾。深入研究发现,在单、双子叶植物分化之后,DREB的碱基组成发生了较大变化,单子叶植物的GC含量和GC3s值比较大,部分双子叶植物的GC3s值大于0.5,符合双子叶植物GC3s值通常大于0.5的规律[22],而有些双子叶植物的GC3s值小于0.5,可能是由于单子叶植物向双子叶植物进化过程中,GC到AT突变压力比AT到GC突变压力高,这一效应同时会反映在同义密码子的第3位碱基上,导致密码子的第3位不是A就是T,反之亦然[23]。 聚类分析和密码子偏好性分析结果存在一定差异,但都可以将单、双子叶植物分别归类,而且将属于禾本科、藜科的不同物种优先聚类在一起。与植物传统分类相比,基于密码子使用偏性的聚类分析可能反映了DREB特殊的进化规律,在一定程度上反映基因特殊的个体进化规律[24],可能是物种进化关系分析中对系统发育分析的重要补充。而基于DREB的CDS序列系统进化树正确反映了物种间的进化关系,在一定程度上优于密码子使用频率的聚类树状图[25],与传统植物分类学结果更接近。利用密码子使用偏性进行聚类分析结合基于CDS序列和基于密码子使用偏性相结合的分析有助于提高分析结果的准确度,更加客观地说明物种的进化进程。 基因的表达效率与受体系统密码子使用偏性息息相关。根据受体生物密码子使用偏好性,对目标基因进行序列改造和修饰,是提高表达效率的有效途径之一[26]。通过比较常用异源表达受体、模式植物与小麦DREB密码子使用频率发现,与小麦密码子使用频率差异更小是酵母菌,更适合做小麦DREB的表达受体,但是密码子使用频率差异的存在预示在做异源表达时需要对小麦DREB基因进行适当的改造。小麦DREB基因与烟草和番茄基因组密码子使用频率差异较大,在遗传转化过程中可能需要更进一步的序列改造和优化;与拟南芥基因组密码子使用频率差异较小,表明拟南芥更适宜作为遗传转化的受体。 本研究利用生物信息学的手段,明确了小麦DREB基因密码子的使用偏好特性,并找到了较合适的异源表达受体。下一步还需适当改造和优化DREB基因,以提高DREB基因在转基因工程研究中表达效率。1.4 DREB基因的聚类分析
2 结果与分析
2.1 小麦DREB密码子的偏性分析

2.2 不同物种的DREB密码子选用偏好性分析

2.3 小麦DREB受体系统的选择



3 讨 论
