不同生长基质的苔藓植物优势种生态位与种间联结
2019-02-21郑越月敖艳艳
刘 艳,郑越月,敖艳艳
重庆师范大学生命科学学院, 重庆 401331
生态位是生态学的重要概念和理论之一,对于理解群落结构和功能、群落内种间关系、生物多样性、群落动态演替和种群进化等方面具有重要作用。种间联结是不同物种在空间分布上的相互关联性,是群落重要的数量和结构特征之一[1]。因此,研究物种生态位和种间联结有助于了解物种对资源的利用能力、物种间协作和竞争关系以及群落演替规律。
苔藓植物是以孢子传播的植物类群之一,从赤道至两极均有分布,能够在土壤、岩石、树干、腐木等不同基质上生长,是森林生态系统中的重要植被组成,在养分循环、水土保持等方面具有重要作用[2-3]。目前关于苔藓植物生态位的研究主要见于温带森林生态系统[4-6]和城市生态系统[7-10],而较少聚焦亚热带森林生态系统。另一方面,关于苔藓植物种间联结的定量研究较少[11-12]。
重庆大巴山国家级自然保护区(下简称大巴山保护区)是典型的北亚热带森林生态系统,属于我国35个生物多样性优先保护区域之一的大巴山区,具有较高的科学研究价值,记录有苔藓植物57科141属390种[13]。本文通过对该保护区内岩石和土壤两种生长基质上的苔藓植物群落进行样方调查,分别计算物种生态位宽度、生态位重叠值和种间联结系数,探究以下问题:(1)不同生长基质上苔藓植物优势种的组成及其对资源的利用能力和种间关系;(2)石生和土生苔藓群落的演替阶段,以期为揭示苔藓植物对环境的适应性和种间关系提供科学依据。
1 研究地区与研究方法
1.1 研究区概况
大巴山保护区(31°37′27″—32°12′15″ N,108°27′07″—109°16′40″ E)位于重庆市东北部的城口县境内。该地区属亚热带季风气候,年均气温13.8℃,年均降水量1261.4 mm,年均日照时数1534 h,年无霜期234 d,年平均相对湿度78%。保护区内海拔落差较大(754—2685 m),包括13个植被类型[14]。
1.2 研究方法
2017年5—6月和8月,先后在大巴山保护区内开展样方调查。样地设置主要从不同生长基质、不同海拔区间和不同乡镇几个方面进行综合考虑。基于前期对大巴山保护区苔藓植物的区系调查结果[13],发现岩石基质的种类占该地区总种数的86.67%,而土壤基质的种类仅占总种数的19.23%,表明该地区石生苔藓植物占绝对优势。最终,在海拔800—2400 m区间分别设立石生苔藓植物样地40个和土生样地19个,包括9个植被类型。59个样地在9个植被类型中的海拔信息如下:常绿阔叶林990—1554 m、落叶阔叶林960—2192 m、常绿落叶阔叶混交林1030—1341 m、常绿阔叶灌丛827—1445 m、落叶阔叶灌丛1120—2245 m、灌草丛2260 m、常绿针叶灌丛2345 m、竹林1680—2254 m、沼泽2275 m。两种生长基质下,各植被类型所调查的苔藓植物群落数目见表1。每个样地内,间隔2 m随机设置大小为20 cm×20 cm的样方20个,共调查样方1180个。

表1 不同植被类型下岩石和土壤基质的苔藓植物样地数
每个样方中不同种类的盖度用划分为16个5 cm×5 cm的网格进行测定。样方中所有苔藓植物带回实验室鉴定到种。种类鉴定和学名主要参考《中国苔藓志》[15-21]。凭证标本保存在重庆师范大学生物标本馆。
1.3 数据处理
1.3.1 物种重要值
根据下式计算苔藓植物重要值:
重要值=(相对盖度+相对频度)/2
式中,相对盖度=(某种苔藓植物样地内盖度/样地内所有苔藓植物盖度之和)×100%,相对频度=(某种苔藓植物样地内频度/样地内所有苔藓植物频度之和)×100%。
1.3.2 生态位宽度
选取重要值为资源利用的参数,将不同植被类型作为资源状态,采用Shannon-Wiener指数[22]计算生态位宽度,公式如下:
式中,Bi为种i的生态位宽度,Pij=nij/Ni,nij为种i在植被类型j上的重要值,Ni为种i在所有植被类型上的重要值之和,Pij为种i在植被类型j上的重要值占该种在所有植被类型上的重要值总和的比例,r为植被类型总数。
1.3.3 生态位重叠
采用Pianka重叠指数[23]计算生态位重叠,公式如下:
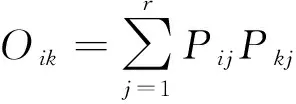
式中,Pij和Pkj分别为种i和种k在植被类型j上的重要值。
1.3.4 种间总体关联性检验
采用方差比率(VR)[24]对种间总体关联程度进行检验,并运用W值检验关联的显著性,公式如下:


1.3.5 种间关联分析
种间关联分析采用卡方检验和计算联结系数相结合的方法进行分析。种间关联的显著程度采用Yates的连续校正公式[1]进行卡方检验,公式如下:
式中,N为样地总数,a为种对同时出现的样地数,b,c为仅种对之一出现的样地数,d为种对均不出现的样地数。当6.635>χ2>3.841,表明两个种相互关联(P<0.05);当χ2>6.635,表明两个种极显著关联(P<0.01)。
种间关联程度通过计算联结系数(AC)测定[1],公式如下:
AC= (ad-bc) /(a+b) (b+d) (ad≥bc)
AC= (ad-bc) /(b+d)(c+d) (bc≥ad,d≥a)
AC= (ad-bc) /(a+b)(a+c) (bc≥ad,d>a)
AC值的范围在[-1, 1],越接近-1,表明种对间负联结程度越高;越接近1,表明种对间正联结程度越高;AC值为0,表明两个物种完全独立。
2 结果与分析
2.1 两种生长基质的优势种在不同植被类型中的重要值
40个石生样地和19个土生样地分别包括154种和77种苔藓植物。选取物种重要值排前15位(约占总物种数的10%)和前10位(占总物种数的13%)的物种分别作为岩石(表2)和土壤(表3)两种生长基质的优势种。石生苔藓优势种的重要值在落叶阔叶林中最高,其次是在落叶阔叶灌丛中、再次是在常绿阔叶林中(表2);土生苔藓优势种的重要值在落叶阔叶林中最高,其次是在竹林中,再次是在落叶阔叶灌丛中(表3)。

表2 石生苔藓植物优势种在不同植被类型中的重要值和生态位宽度
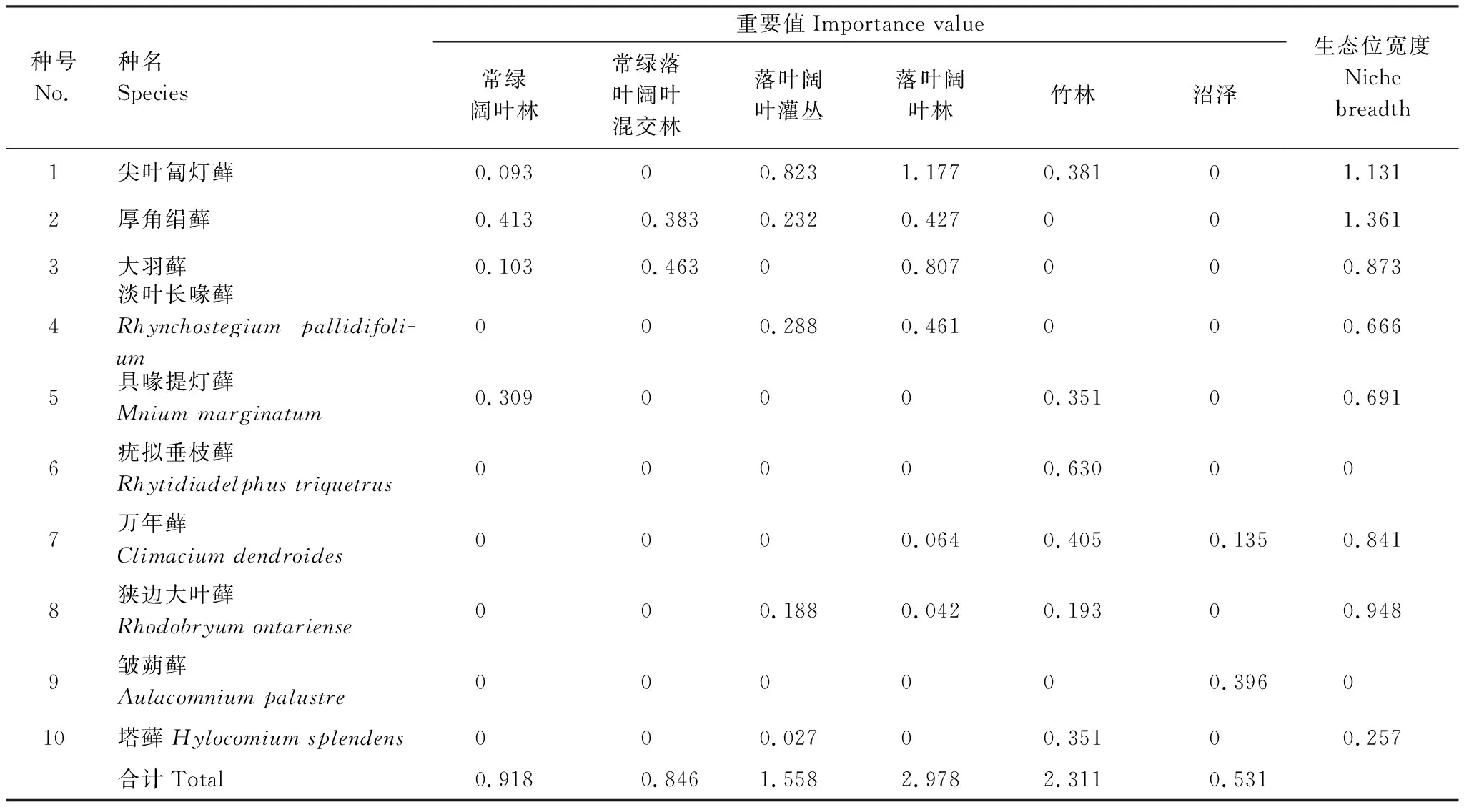
表3 土生苔藓植物优势种在不同植被类型中的重要值和生态位宽度
2.2 生态位宽度
从表2可知,15种石生苔藓植物优势种的生态位宽度为0.574—1.783,平均值为1.141。其中大羽藓Thuidiumcymbifolium的生态位宽度最大,分布范围最广。除灌草丛外,在7个植被类型中均有分布(表2)。相反,匐枝青藓Brachytheciumprocumbens的生态位宽度最窄,仅在两个植被类型中有分布。从表3可知,10种土生苔藓植物优势种的生态位宽度为0—1.361,平均值为0.677。其中厚角绢藓Entodonconcinnus的生态位宽度最宽,在4个植被类型中均有分布,而疣拟垂枝藓Rhytidiadelphustriquetrus和皱蒴藓Aulacomniumpalustre的生态位宽度最窄,各自仅在1个植被类型中有分布。
2.3 生态位重叠
对15种石生苔藓植物优势种的生态位重叠进行计算,共计105对种对(表4)。生态位重叠值介于[0.048, 0.939],平均值为0.535,主要集中在[0.5, 0.8],占种对总数的40.00%。其次,在[0.2, 0.5]的占30.48%。牛角藓Cratoneuronfilicinum与溪边青藓Brachytheciumrivulare的生态位重叠值最大,表明它们利用的资源状态相似性非常高。
对10种土生苔藓植物优势种的生态位重叠进行计算,共计45对种对(表5)。生态位重叠值介于[0, 0.998],平均值为0.360,主要集中在小于0.2的区间,占种对总数的44.44%。其次,在[0.5, 0.8]的占26.67%。有13对种对的生态位重叠值为0,主要是生长在沼泽的皱蒴藓与其他土生苔藓植物优势种,表明这些种对的两个物种间完全没有重叠,利用不同的资源状态。相反,疣拟垂枝藓与塔藓Hylocomiumsplendens的生态位重叠值最大,表明它们利用的资源状态几乎完全相同。
2.4 种间联结性
岩石(VR=0.064/0.392)和土壤(VR=0.110/0.442)两种基质的苔藓优势种种间总体联结性的方差比率VR均小于1,表明种间总体上为负联结。分别计算统计量W检验VR的显著性,石生苔藓的W=6.519,不在置信区间(26.51, 55.76)内;土生苔藓的W=4.718,也不在置信区间(10.12, 30.14)内,表明物种间联结性显著。
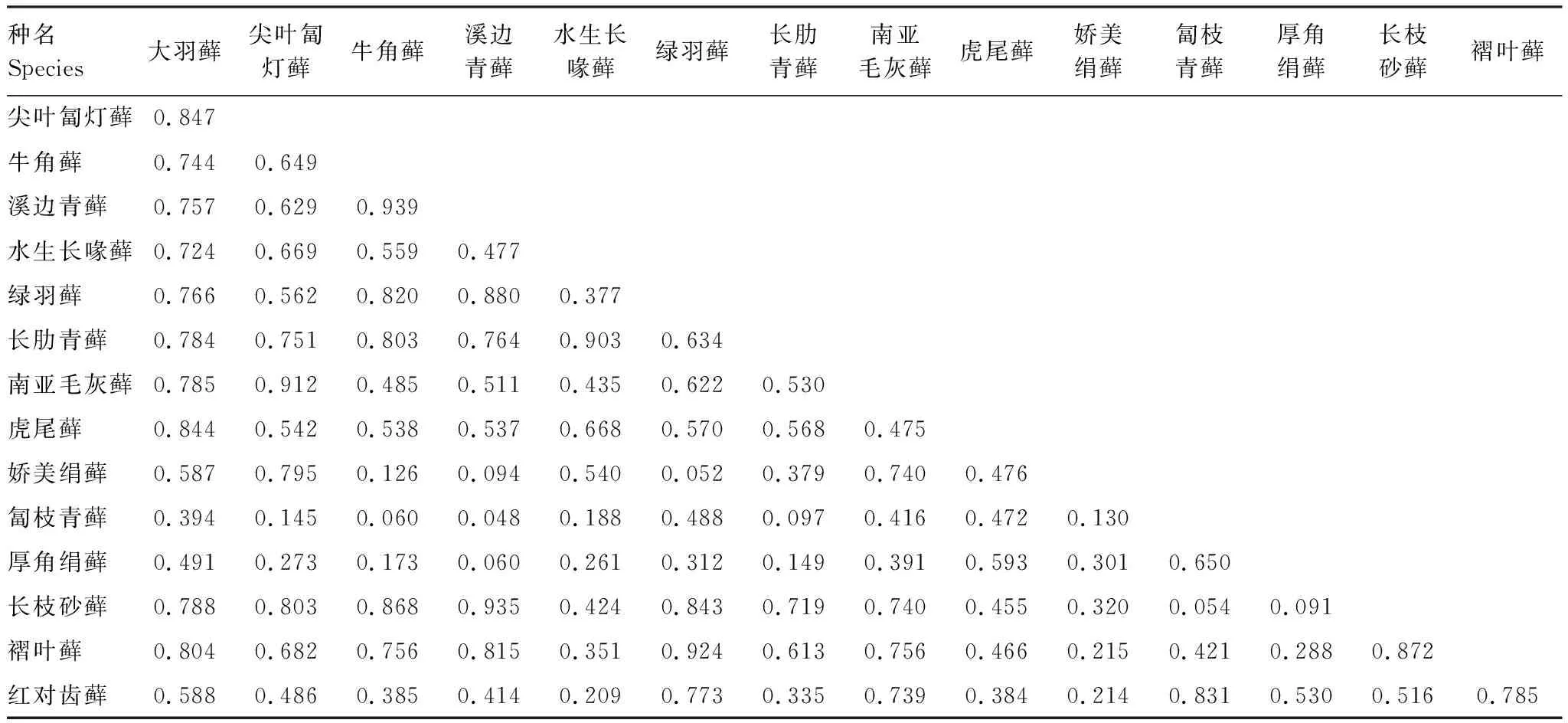
表4 石生苔藓植物优势种生态位重叠值
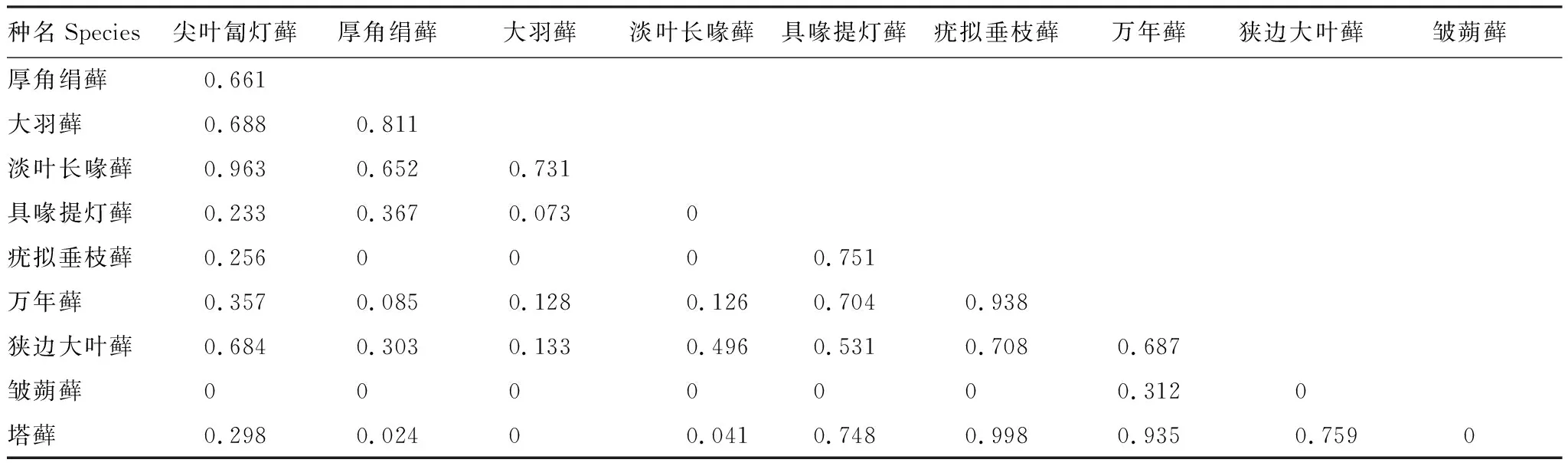
表5 土生苔藓植物优势种生态位重叠值
15种石生苔藓植物优势种形成的105对种对中,联结系数AC值范围在>-0.01、[-0.34, -0.01]、[-0.67, -0.34]和<-0.67,分别占总种对数的26.67%、14.29%、6.67%和52.38%。有43对种对的AC值为-1,表明负联结程度最高,主要是匐枝青藓、长枝砂藓Racomitriumericoides和褶叶藓Palamocladiumnilgheriense与其他石生优势种形成的种对。总体上看,种间负联结有77对,占总种对数的73.33%,其中8对表现出极显著性,18对表现出显著性;种间正联结有28对,占总种对数的26.67%,其中4对表现出极显著性,7对表现出显著性(图1)。
10种土生苔藓植物优势种形成的45对种对中,联结系数AC值范围在<-0.69、 [-0.69, -0.38]、[-0.38, -0.07]和>-0.07,分别占总种对数的57.78%、4.44%、8.89%和28.89%。有23对种对的AC值为-1,表明负联结程度最高,主要是皱蒴藓和厚角绢藓与其他土生优势种形成的种对。总体上看,种间负联结有25对,占总种对数的55.56%,其中7对表现出极显著性,12对表现出显著性;种间正联结有20对,占总种对数的44.44%,其中1对表现出显著性(图2)。
2.5 同一物种的生态位和同一种对种间联结性比较
在岩石和土壤两种生长基质上,大羽藓、尖叶匐灯藓Plagiomniumacutum和厚角绢藓均为优势种之一。比较其生态位宽度发现,这3种藓类在岩石基质的生态位宽度比其在土壤基质的更大(表2和表3)。比较同一种对在两种生长基质中的生态位重叠值发现(表4和表5),大羽藓-尖叶匐灯藓种对在岩石基质的生态位重叠值(0.847)高于其在土壤基质的生态位重叠值(0.688);而大羽藓-厚角绢藓、尖叶匐灯藓-厚角绢藓两个种对在岩石基质的生态位重叠值(0.491和0.273)均小于其在土壤基质的生态位重叠值(0.811和0.661)。
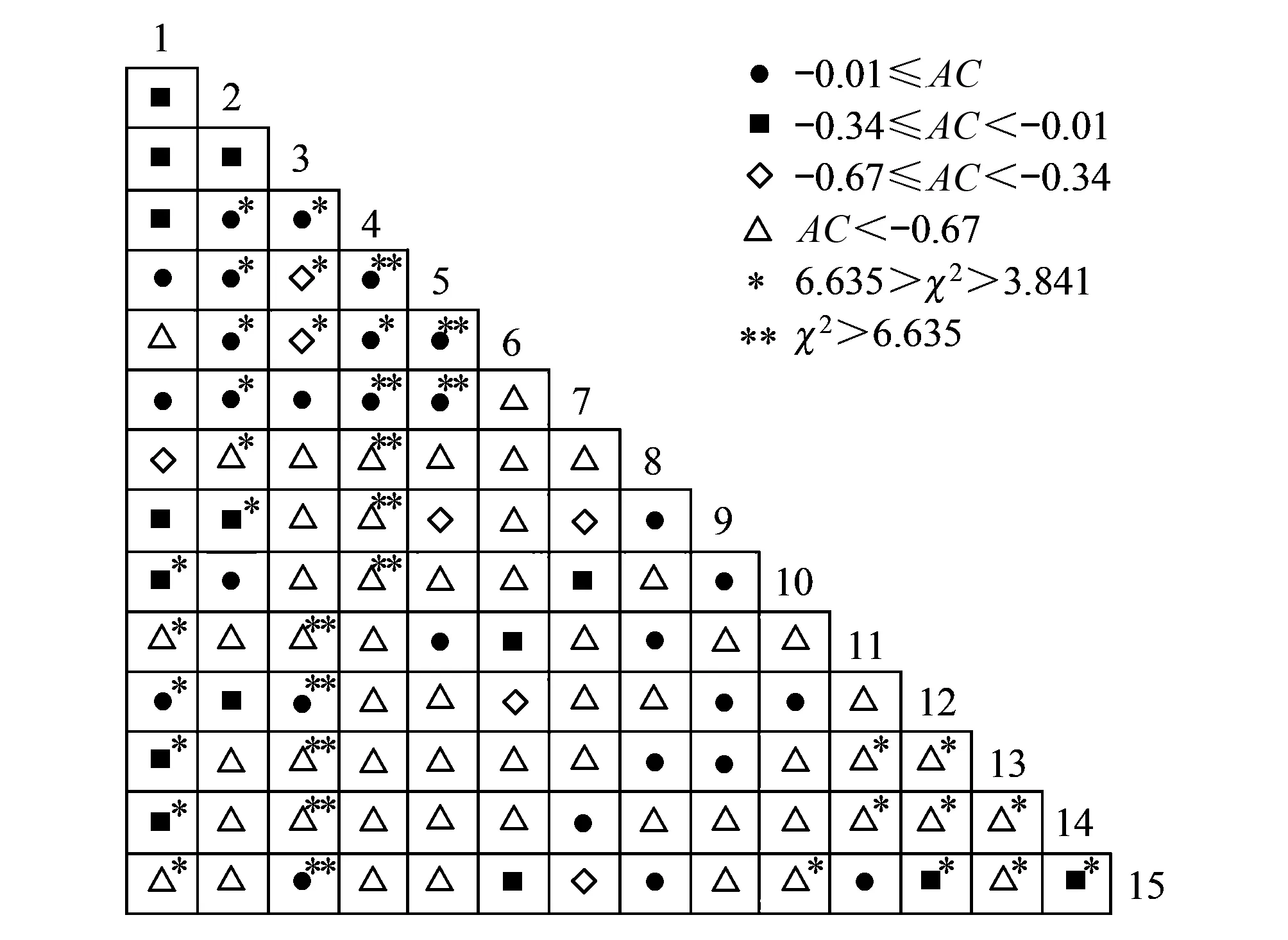
图1 石生苔藓植物优势种种间联结性半矩阵图Fig.1 Semi-matrix of interspecific association of dominant bryophytes on rocksAC:联结系数, association coefficients; 1:大羽藓Thuidium cymbifolium, 2:尖叶匐灯藓Plagiomnium acutum, 3:牛角藓Cratoneuron filicinum, 4:溪边青藓Brachythecium rivulare, 5:水生长喙藓Rhynchostegium riparioides, 6:绿羽藓Thuidium assimile, 7:长肋青藓Brachythecium populeum, 8:南亚毛灰藓Homomallium simlaense, 9:虎尾藓Hedwigia ciliata, 10:娇美绢藓Entodon pulchellus, 11:匐枝青藓Brachythecium procumbens, 12:厚角绢藓Entodon concinnus, 13:长枝砂藓Racomitrium ericoides, 14:褶叶藓Palamocladium nilgheriense, 15:红对齿藓Didymodon asperifolius
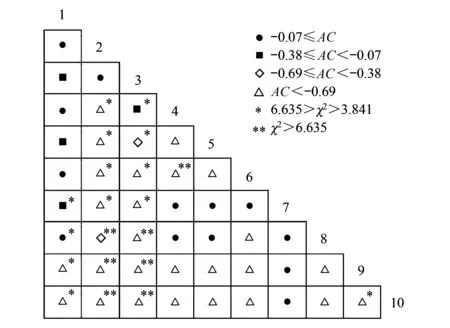
图2 土生苔藓植物优势种种间联结性半矩阵图Fig.2 Semi-matrix of interspecific association of dominant bryophytes on soil1:尖叶匐灯藓, 2:厚角绢藓, 3:大羽藓, 4: 淡叶长喙藓Rhynchostegium pallidifolium, 5: 具喙提灯藓Mnium marginatum, 6: 疣拟垂枝藓Rhytidiadelphus triquetrus, 7: 万年藓Climacium dendroides, 8: 狭边大叶藓Rhodobryum ontariense, 9: 皱蒴藓Aulacomnium palustre, 10: 塔藓Hylocomium splendens
比较种对间的联结系数AC发现,大羽藓-尖叶匐灯藓种对在两种基质中均表现出负联结,而大羽藓-厚角绢藓种对均表现出正联结。尖叶匐灯藓-厚角绢藓种对在岩石基质中表现出负联结,而在土壤基质中表现出正联结(图1和图2)。
3 讨论
3.1 苔藓植物生态位
生态位宽度反映物种对环境的适应性。物种生态位宽度越宽,适应环境、利用资源的能力就越强。同样,在大巴山保护区,一些苔藓植物生态位宽度较大,分布范围较广,例如大羽藓、尖叶匐灯藓,在所调查的绝大多数植被类型中均有分布。相反,一些苔藓植物生态位宽度较小,例如疣拟垂枝藓和皱蒴藓的生态位宽度为0(表3),是因为它们分别只在大巴山保护区内的亚高山竹林和亚高山沼泽中有分布,并常常大片生长,形成单一的优势群落。
同一物种在不同生长基质上的生态位宽度存在差异。例如,大羽藓、尖叶匐灯藓和厚角绢藓在岩石基质上的生态位宽度均比其在土壤基质更大,表明它们在岩石基质上的适应性更广,进而说明生长基质会影响苔藓植物利用资源的能力。这与郭磊等对小秦岭国家自然保护区苔藓植物属的生态位研究结果一致[6]。
生态位重叠值大小反映物种之间利用资源的相似程度[25]。研究显示,苔藓植物[6]生态位宽度越大,与其他种的生态位重叠值也会较大。本研究结果支持上述观点。在大巴山保护区,石生苔藓优势种的平均生态位(1.141)明显高于土生苔藓优势种(0.677),而前者种对间生态位平均重叠值(0.535)也明显大于后者(0.360)。
3.2 苔藓植物种间联结与群落演替
理论上认为,群落结构和种类组成越稳定,种间关系趋于正相关[26]。方差比率VR和统计量W检验结果显示,石生和土生苔藓植物种间总体上均为显著负联结,表明群落处于演替的前期或中期。这一结果一方面可能与苔藓植物本身的生物学特性有关,易受环境因素的影响[4,12]。在大巴山保护区,苔藓群落主要受农业生产生活和旅游开发的影响。另一方面,与所调查区域的植被大环境有关。处于顶级群落的原生林由于历史上的过度采伐,面积相对较小。
本研究结果显示,一些小生境类似的优势种表现出明显负联结。例如,牛角藓和水生长喙藓Rhynchostegiumriparioides多生长在溪流中(边)的(流水)岩石上。然而它们相伴出现的概率很小(7.5%),表现出显著负联结(图1)。又例如,属于近缘种的疣拟垂枝藓和塔藓通常分布在大巴山保护区的亚高山竹林下,生态位重叠值非常高(0.998)(表5)。然而它们并没有出现在同一样地,表现出极显著负联结(图2)。这与若两物种对环境要求相似,彼此间表现出正联结的观点[26-27]不一致。这可能是因为上述种对在资源有限的环境中,存在种间竞争或排它作用导致。相反,在泥炭地常见的泥炭藓Sphagnumpalustre与皱蒴藓表现出极显著正联结。这是因为两物种间有利他作用存在[11]。需要指出的是,计算种间联结系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间联结性产生影响[26]。例如,研究表明,样方大小对泥炭地苔藓植物种间联结的影响较大[11]。因此,种间关联的内在机制还需要进一步从植物生理、遗传学、分子生态学等多角度开展研究才能合理解释。
