盐胁迫下野牛草相连分株根尖钙离子流变化
2019-02-19钱永强孙振元
罗 迪,罗 栋,2,钱永强,孙振元
(1. 中国林业科学研究院林业研究所 / 森林培育国家林业局重点实验室,北京 100091;2. 广东省深圳市福田区城市管理局,广东 深圳 518036))
克隆植物由各种等级的构件组成,各个构件通过连接物和间隔子进行连接,形成一个形态和生理上相连的克隆系统[1]。自然生境中资源常呈异质性分布,表现为浓度、尺度、格局和对比度上的差异[2-4]。为了适应异质生境,克隆植物通过节间连接,使光合同化物、水分、营养甚至信号分子在各克隆构件间运输和共享,即生理整合,并通过在不同部位表型可塑性修饰[5-8],实现物质与能量的最优分配[9-10],从而增强了克隆植物在栖息地中的竞争力和入侵能力[11-14]。因此,与非克隆植物相比,克隆植物表现出更强的空间拓展和生态适应能力[15-17]。
生理整合是克隆植物区别于非克隆植物最主要的特征之一[18-19]。生理整合的发生是以感知异质胞外信号为前提的。因此,探索克隆植物相连分株对异质生境的应答及适应性机制,对解析克隆植物极强的生态适应性、挖掘克隆植物的应用潜力具有重要意义。野牛草(Buchloe dactyloides )是一种匍匐茎型克隆植物,可通过快速克隆生长,形成许多相对独立或潜在独立的克隆分株,组成不同等级的克隆构件,连成一个形态和生理上相连的克隆分株系统[20-21],是研究克隆植物对异质生境响应、生理整合以及对克隆植物表型可塑性影响的理想材料。
众多研究表明,Ca2+是植物生长发育必需的营养元素,同时Ca2+作为重要的第2信使,通过胞内钙库Ca2+的释放及胞间Ca2+振荡等时空动态变化,作为信号物质的同时还可以结合细胞内Ca2+受体钙调蛋白(CaM)起作用,参与应答多种非生物胁迫,并发挥着重要的信号转导调控机制[22-24]。
基于此,本研究对相连野牛草克隆分株设置了异质性NaCl胁迫,利用非损伤微测技术测定正常条件和NaCl胁迫下野牛草克隆片段姊妹株根尖Ca2+净流速量的方向和变化规律,研究野牛草分株根尖Ca2+流对NaCl胁迫的响应及生理整合对相连分株根尖Ca2+流的调控作用,进而为更深入地了解克隆植物对异质生境的表型可塑性提供理论依据。
1 材料与方法
1.1 材料培养
野牛草通过克隆生长形成许多相对独立的克隆分株,同一匍匐茎上可以先后形成多个克隆分株,通过间隔子相连的野牛草克隆分株形成一个相对独立的克隆片段。选取长势良好的野牛草单株培养于中国林业科学研究院科研温室中以获得大量匍匐茎。将匍匐茎节点浸于纯净水中24 h,使其生根,水量以刚没过匍匐茎为准。选取野牛草匍匐茎顶端开始数的第3株和第4株长势一致、根长一致、生长成熟的相连克隆片段作为试验材料。将克隆片段中较年轻的植株定义为妹株(第3株),较老的植株(第4株)定义为姊株。

图1 野牛草克隆分株的NaCl处理示意图Figure 1 Buchloe dactyloides clonal fragments under NaCl stress
1.2 处理方法
1.2.1 NaCl处理
将克隆片段的相连姊妹株分别放置于两个相邻的、直径为30 mm培养皿中,按处理每皿中加入20 mL相应的含NaCl或不含NaCl测试液,平衡30 min后测定离子流的变化,稳定后记录。NaCl处理浓度为 200 mmol·L-1。共设置 4 个处理 (图 1),分别为,对克隆片段姊株进行NaCl胁迫处理(T1);对克隆片段妹株进行NaCl胁迫处理(T2);对克隆片段姊株和妹株同时进行NaCl胁迫处理(T3);对照不进行NaCl胁迫处理(TCK)。每处理6个重复,分别测定姊株和妹株根尖Ca2+离子流。
1.2.2 LaCl3处理
将克隆片段的相连姊妹株分别放置于两个相邻的、直径为30 mm培养皿中,按处理每皿中加入20 mL相应的含NaCl或不含NaCl测试液。NaCl处理浓度为 200 mmol·L-1,LaCl3处 理 浓 度 为 20 mmol·L-1,每处理6个重复,设置4个处理(图2)。分别为,用LaCl3处理相连克隆片段姊株30 min,对姊株进行NaCl胁迫处理(TL1);用LaCl3处理相连克隆片段妹株30 min,对妹株进行NaCl胁迫处理(TL2);用LaCl3处理相连克隆片段的姊株和妹株30 min,对姊株和妹株同时进行NaCl胁迫处理(TL3);用LaCl3处理相连克隆片段姊株 30 min,不进行NaCl胁迫处理(TLCK)。处理后分别测定姊株和妹株根尖Ca2+离子流。
图2 野牛草克隆分株的NaCl和LaCl3处理示意图Figure 2 Buchloe dactyloides clonal fragments under NaCl stress and LaCl3 treatment
1.3 离子流测定
采用非损伤微测系统(Bio-IM NMT,美国扬格公司)测定野牛草分株根系Ca2+离子流变化。测定部位为根尖分生区,距离根部顶端250 μm处。Ca2+测试液为 0.1 mmol·L-1CaCl2、0.1 mmol·L-1KCl、0.1 mmol·L-1MgCl2、0.5 mmol·L-1NaCl、0.2mmol·L-1Na2SO4、0.3 mmol·L-1MES,pH 6.0;盐胁迫处理时 测 试 液 中 再 加 入 200 mmol·L-1NaCl。 低 浓度(0.05 mmol·L-1)Ca2+校 正 液 为 0.05 mmol·L-1CaCl2、0.1 mmol·L-1KCl、0.1 mmol·L-1MgCl2、0.5mmol·L-1NaCl、0.2 mmol·L-1Na2SO4、0.3 mmol·L-1MES,pH 6.5;高浓度 (0.5 mmol·L-1)Ca2+校正液为 0.5 mmol·L-1CaCl2、 0.1 mmol·L-1KCl 、 0.1 mmol·L-1MgCl2、0.5 mmol·L-1NaCl、0.2 mmol·L-1Na2SO4、0.3 mmol·L-1MES,pH 5.5。
将测定的数据代入Fick第一扩散定律公式,计算 Ca2+流动速率[单位为 pmol·(cm2·s)-1]。公式如下:
J = -D·dc/dx。
式中:J为电极移动方向的离子流;dx为电极移动距离,dc为电极移动的两点间的离子浓度差,D为测定离子的扩散常数。离子流速计算结果为正值时表明Ca2+流出细胞(外流);计算结果为负值时表明Ca2+流入细胞(内流)。
1.4 数据分析
利用单因素方差分析方法对各处理间姊株和妹株的Ca2+流差异进行分析,并用Duncan检验进行多重比较;利用独立样本T检验的方法比较相同处理下姊株和妹株间的Ca2+流差异;采用SPSS 16.0(SPSS Inc., Chicago, IL, USA)统计软件进行分析。
2 结果
2.1 姊妹克隆分株根尖Ca2+流对NaCl胁迫的响应
各处理姊株根尖Ca2+净流量均呈现外流的状态(图3)。与对照(CK)相比,NaCl胁迫下(T1、T2和T3)姊株根尖的 Ca2+净外流量显著增加 (P<0.05),但各NaCl胁迫处理间姊株根尖Ca2+净流量并无显著差异 (P > 0.05)。
与姊株根尖离子流结果相似,各处理妹株根尖的Ca2+净流量也均呈现外流的状态(图3)。与对照(CK)相比,单独对妹株进行NaCl胁迫(T2)和同时对姊妹株进行NaCl胁迫时(T3)显著提高了妹株根尖 Ca2+净外流量 (P<0.05)。而单独对姊株进行NaCl胁迫时(T1),妹株根尖Ca2+净流量并无明显变化 (P > 0.05)(图 4)。

图3 野牛草姊株和妹株根尖Ca2+净流量对NaCl胁迫的响应Figure 3 Ca2+ net flux responses of older and younger ramet root tips of Buchloe dactyloidesunder different NaCl treatments不同小写字母表示不同处理间差异显著(P<0.05)。图5同。Different lowercase letters indicate significant differences among different treatments at the 0.05 level; similarly for Figure 5.
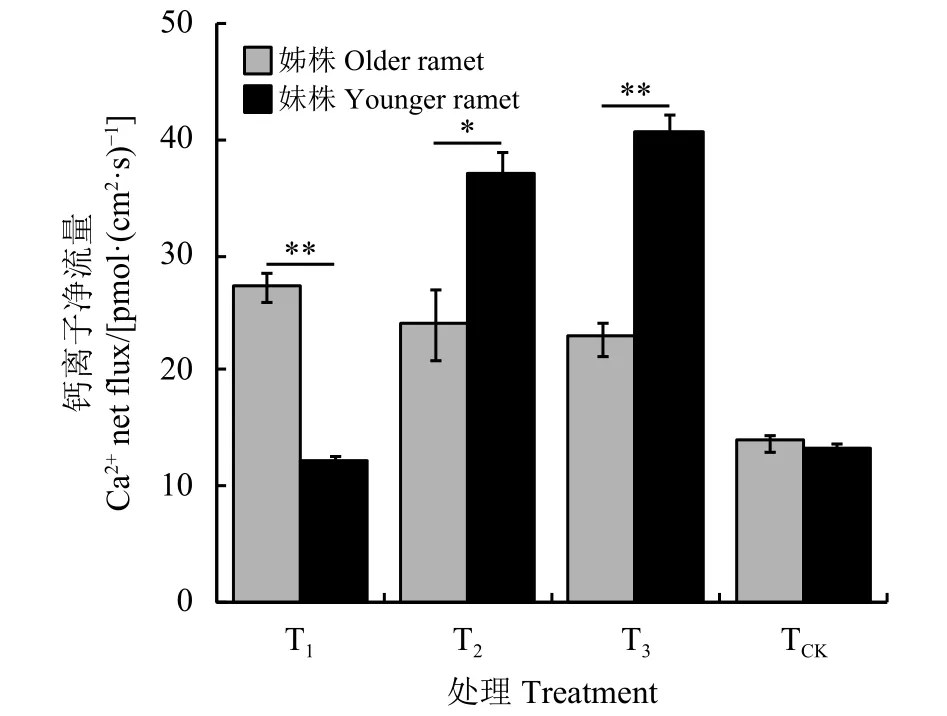
图4 野牛草姊妹株根尖Ca2+净流速在相同NaCl胁迫下的差异Figure 4 Differences between Ca2+ net flux of older and younger ramet root tips of Buchloe dactyloides under the same NaCl stress*和**分别表示相同处理下姊妹株间差异显著(P<0.05)或极显著 (P<0.01)。图 6 同。* and ** indicate significant differences between older and younger ramets under the same treatment at 0.05 and 0.01 levels, respectively;similarly for Figure 6.
2.2 相同NaCl胁迫下姊株和妹株根尖Ca2+流比较
对照(TCK)的姊株和妹株根尖Ca2+净流量无显著差异 (P > 0.05)(图 5)。单独对姊株进行 NaCl胁迫时(T1),姊株根尖Ca2+净外流量显著高于妹株(P <0.05)。单独对妹株进行NaCl胁迫时(T2),妹株根尖Ca2+净外流量显著高于姊株(P<0.05)。对姊株和妹株同时进行NaCl胁迫时(T3),妹株根尖Ca2+净外流量显著高于姊株(P<0.05)。
2.3 姊妹克隆分株根尖Ca2+流对LaCl3的响应
LaCl3处理后,姊株根尖Ca2+仍呈现外流状态,各处理间差异并未达到显著水平 (P > 0.05)(图 6)。LaCl3处理后,妹株根尖Ca2+仍呈现外流状态,各处理间差异也并未达到显著水平(P > 0.05)(图5)。加入LaCl3后,姊株和妹株根尖Ca2+外流均增加,且与未加LaCl3处理对比差异达到极显著水平 (P<0.01)(图 6)。
3 讨论与结论
非损伤微测技术 (non-invasive micro-test technique,NMT)也称无损微测技术,是一种选择性离子和分子检测技术[25-27]。它利用特异性离子或者分子选择性电极,在不损伤样品的基础上,测定多种离子和分子进出活体样品的流速、流量和流向,且具有极高的精准度。非损伤微测技术在直接获取组织或细胞外部分子和离子的信息方面具有明显优势,目前已被广泛应用于生命科学的诸多领域[27]。
研究表明NaCl胁迫会引起植物细胞Ca2+浓度的升高[28]。植物细胞质膜和内膜系统上的钙离子通道调控NaCl胁迫诱导Ca2+浓度升高。本研究中,野牛草姊株和妹株根尖Ca2+净流量在所有处理中均呈现外流状态,Ca2+外流主要通过位于质膜上的电压依赖型钙泵完成。即当膜电位去极化或超极化时,钙泵被激活,Ca2+外流[28]。
NaCl胁迫会导致克隆植物的不同构件对资源的吸收产生差异,从而使分株间形成资源的生理整合[15]。生理整合的发生以感知生境异质信号为前提,而生理整合所引起的表型变化也是以信号调控为基础的。野牛草克隆分株可以感知NaCl胁迫信号,引起克隆分株根尖Ca2+发生变化。不同生理整合条件下的克隆分株根尖Ca2+流存在差异。生长在正常营养条件下的姊株和妹株根尖Ca2+外流流速差异不明显。当对姊株进行NaCl胁迫时,姊株感受到胁迫,Ca2+外流迅速增加,根据“收益-成本”原则[29],姊株并没有马上将胁迫信号传递给妹株,因而妹株Ca2+外流变化不明显。当对妹株进行NaCl胁迫时,妹株Ca2+外流迅速增加,同时妹株将胁迫信号传递给姊株,共同对胁迫作出响应,姊株Ca2+外流也迅速增加,妹株Ca2+流速显著高于姊株。当姊株和妹株均受到NaCl胁迫时,姊株和妹株Ca2+外流均迅速增加,且妹株Ca2+流速显著高于姊株。野牛草姊妹株间存在NaCl信号的整合,调控了NaCl胁迫下野牛草分株根尖Ca2+流的变化。
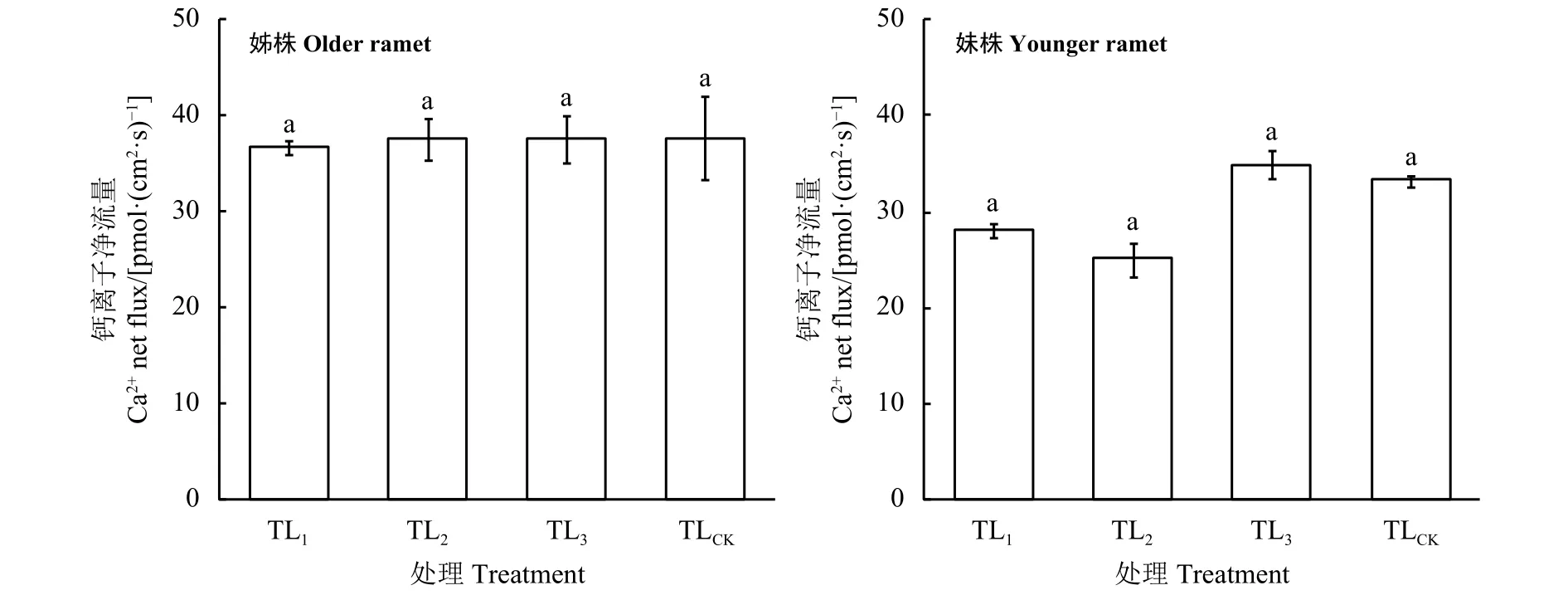
图5 野牛草根尖Ca2+净流速对LaCl3胁迫的响应Figure 5 Ca2+ net flux responses of Buchloe dactyloides root tips to different LaCl3 treatments
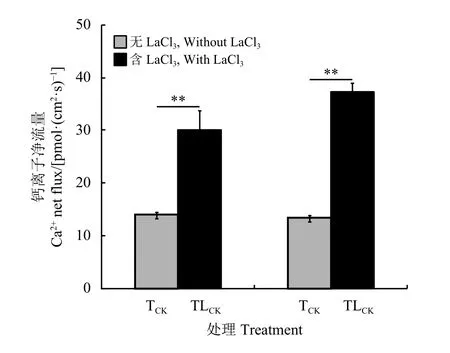
图6 LaCl3对野牛草根尖Ca2+净流速的影响Figure 6 Effects of LaCl3 on Ca2+ net flux of Buchloe dactyloides root tips
LaCl3阻断了Ca2+通道,阻止分株间Ca2+的运输。存在营养生理整合的克隆分株用LaCl3处理后虽然会引起根尖Ca2+外流增加,但各NaCl胁迫处理下Ca2+外流与对照相比差异均不显著。LaCl3阻断了分株间Ca2+的运输,NaCl胁迫信号不能被克隆分株根尖所感知并在分株间转导。这证明,Ca2+参与克隆分株对异质营养的感知和应答,调控了克隆分株间的营养生理整合。
