一株同步硝化-反硝化菌的絮凝特性
2019-02-15梁锡宏李政威吴重德周荣清
李 丹,梁锡宏,李政威,金 垚,吴重德,周荣清
(四川大学 轻纺与食品学院 皮革化学与工程教育部重点实验室,四川 成都 610065)
目前,最常用的脱氮方法是在好氧条件下利用自养菌进行硝化,在厌氧条件下利用异养菌进行反硝化[1]。然而,独立的硝化反硝化池占地面积大、耗时长、花费高,且操作不便[2]。同时,由于自养硝化菌生长缓慢,易受高浓度的氨、有机物和其他化学元素的影响,使得硝化成为了脱氮过程的限速步骤[3]。近年来,研究人员从环境中分离获得了异养硝化-反硝化菌[4-6],这些异养菌比自养菌生长更快,能够利用有机物作为碳源,同时在好氧条件下将含氮物质转变成N2。在同一个反应器中进行硝化-反硝化可以降低操作费用,提高氮的去除率,而对同步异养硝化-好氧反硝化菌研究的不断深入,能为更好地实现同步硝化-反硝化提供更多的理论和实践依据[7]。
活性污泥胞外聚合物(EPS)来源于细胞的分泌溶解、大分子水解以及废水中吸附的有机物[8],对活性污泥的形成、污泥絮体结构、脱水沉降性能以及重金属的吸附具有重要的影响[9]。据报道,EPS还会与水体中的各种元素发生生物、化学和物理反应,如配位、吸附、吸收和絮凝反应等,对研究水体与生物之间物质输送和交换的途径具有重要影响[10]。此外,胞外聚合物还能提高微生物对重金属的抗性和吸附[11]。目前,对菌体胞外聚合物的研究多集中在人体口腔微生物[12]和益生菌等肠道微生物[13],而针对硝化-反硝化菌EPS的研究很少。此外,硝化反硝化菌EPS是硝化池和反硝化池中活性污泥能否形成絮凝体的关键。
本文中,笔者研究了1株脱氮菌的硝化反硝化性能和自聚集性能,并对其产生絮凝性的胞外聚合物特性进行研究,以期为未来的硝化反硝化菌胞外聚合物研究提供重要参考。
1 材料与方法
1.1 菌株
克雷伯氏菌(Klebsiellasp.TN-10),从制革废水中筛选得到,保藏于中国典型培养物保藏中心,保藏编号为CCTCC NO:M2017193。
1.2 培养基及菌种培养
硝化培养基(g/L):丙酮酸钠7、(NH4)2SO40.5、NaCl 1、MgSO40.5、EDTA 0.8、FeSO40.4、K2HPO41;pH 7.0。
反硝化培养基Ⅰ(g/L):丙酮酸钠7、NaNO30.5、NaCl 1、MgSO40.5、EDTA 0.8、FeSO40.4、K2HPO41;pH 7.0。
反硝化培养基Ⅱ(g/L):丙酮酸钠7、NaNO20.5、NaCl 1、MgSO40.5、EDTA 0.8、FeSO40.4、K2HPO41;pH 7.0。
从甘油管中将菌种TN-10接种到含50 mL培养基的三角瓶中,在150 r/min、30 ℃条件下培养12 h,将种子液按1%(体积分数)接种量接种到100 mL的上述培养基中,在150 r/min、30 ℃的摇床中进行培养。
1.3 硝化-反硝化能力测定

菌体反硝化性能的测定:将TN-10接种到反硝化培养基Ⅰ和Ⅱ中,培养条件同上,间隔8 h取样,用同样的方式处理样品,测定TN-10在反硝化培养基中的生长和氮的降解情况。
1.4 自聚集性能测定
将活化12 h的种子液按照体积分数1%的接种量接种到硝化培养基中,分别收集培养至对数期、稳定期和衰亡期的发酵液。将样品在10 000 r/min、4 ℃下离心5 min,去除上清液,并用H3PO4缓冲液(PBS)清洗2次,重悬到PBS中,调整菌体OD600至0.6。将30 mL菌悬液放入50 mL离心管中,每间隔1 h取样,测定上清液的OD600,菌体的自聚集性计算可以采用式(1)[14]。
自聚集性=(1-At)/A0×100%
(1)
式中:At为t时刻的OD600;t分别为1、2、3、4和5 h;A0为0时刻的吸光值。
菌体的聚集过程可以看成是可逆的化学反应。使用伪一级动力学方程来描述[15],见式(2)。
At=Ae(1-e-tk1)
(2)
式中:k1=h-1;Ae代表平衡时的自聚集性;At表示t时刻的OD600,与式(1)一致。
1.5 胞外聚合物(EPS)的提取与测定
采用阳离子交换树脂法对菌体胞外聚合物进行提取。将40 mL在硝化培养基中培养12 h的菌体在10 000 r/min、4 ℃下离心5 min,弃上清液,收集菌体。使用超纯水将菌体洗涤2遍并重悬到超纯水中。将细胞悬液放入250 mL三角瓶中并加入一定量的树脂(每克干菌体中加70 g树脂)。将三角瓶放入摇床培养,在150 r/min条件下提取8 h后,离心收集上清液并过0.45 μm滤膜,测定EPS提取液中的蛋白和多糖含量,胞外聚合物的含量以蛋白质和多糖含量表征[16]。多糖的测定采用蒽酮-H2SO4法[17],蛋白质的测定采用改良的BCA法[18]。
1.6 傅里叶红外光谱分析
将EPS提取液使用旋转蒸发仪浓缩,在冷冻干燥仪中进行冷冻干燥,得到干燥的固体结晶。使用傅里叶红外光谱仪对EPS组成进行分析。干燥的EPS与溴化钾按1∶ 100(质量比)混匀,在玛瑙研钵中研磨,并放入压片机进行压片后,用Nicolet380型傅里叶红外光谱仪(美国THERMO Fisher Scientific)对样品进行扫描,分辨率为0.4 cm-1,测定波数范围为4 000~400 cm-1。
1.7 蛋白质二级结构的测定与分析
使用J-715型圆二色光谱仪(JASCO公司)对EPS溶液进行扫描。扫描范围为190~250 nm,在1 mm石英比色皿中以超纯水做空白,每间隔1 nm记录一次数据。使用CD pro程序包中的SELON和CONTIN处理CD谱数据,计算蛋白质二级结构[19]。
2 结果与讨论
2.1 TN-10的硝化-反硝化能力


图1 TN-10的硝化-反硝化性能Fig.1 The nitrifying-denitrifying characteristics of TN-10

2.2 自聚集性能
为了解菌体的自聚集性能,笔者测定了菌体的生长曲线和在不同生长周期的自聚集性能变化情况,结果如图2所示。由图2(a)可知:TN-10从4 h起进入对数期,在24 h到达稳定期,从32 h开始进入衰亡期。收集处于12 h(对数期)、24 h(稳定期)和36 h(衰亡期)的菌体,测定菌体的自聚集率。由图2(b)可知:TN-10表现出较好的自聚集性,其自聚集率达到20%左右,但在不同时期,菌体的自聚集性无显著差异。随着培养时间的延长,菌体的自聚集性呈现缓慢增加的趋势,到达衰亡期,菌体的自聚集率最高。这可能是因为细菌在增殖或衰亡时,会脱落外膜和释放周质蛋白,衰亡期的细胞存在因细胞自融释放的胞内蛋白,使得菌体外的聚合物较对数期和稳定期多[24]。Wang等[16]研究了Enterobactersp.strain FL在不同时间的自聚集性,发现在培养12 h时自聚集率大约为18%,但随着时间的延长,自聚集性能逐渐增加,在培养48 h时,自聚集率能达到50%。由此可见,菌体的自聚集率与菌体的生长周期相关。

图2 TN-10的生长曲线与自聚集性能Fig.2 Growth curve and auto-aggregationability of TN-10
菌体的自聚集动力学表明菌体克服静电排斥后的自聚集能力。本实验中,笔者对菌体的自聚集动力学进行研究,结果如图2(c)所示。由图2(c)可知:不同时期的菌体聚集率随时间的增加而呈现缓慢上升的趋势,但菌体的聚集性在5 h时还未达到完全平衡。同时,从0~5 h,衰亡期菌体的自聚集率始终比对数期和稳定期菌体的自聚集率高,其原因可能是处在不同时期的菌体胞外聚合物性质不同。不同条件下的菌体聚集动力学也相差较大。Zhang等[25]研究活性污泥的自聚集性发现,其自聚集率最大为50%左右,在3 h后保持稳定。
2.3 胞外聚合物组成
为研究菌体呈现出自聚集性能的原因,对菌体的胞外聚合物进行研究。为避免胞内蛋白溢出和细胞外膜脱落对菌体胞外聚合物成分测定的干扰,收集对数期的菌体对其胞外聚合物组成进行研究,结果如表1所示。由于胞外聚合物的主要成分为蛋白质、多糖等,测定蛋白质和多糖的含量来表征EPS[26]。由表1可知:蛋白质含量为每克干细胞中23.84 mg(相当于15.11 mg/L),多糖的含量为每克干细胞中18.64 mg(11.81 mg/L),胞外聚合物以蛋白质和多糖的总量计为每克干细胞中42.48 mg(26.81 mg/L)。TN-10的EPS中的蛋白质含量高于多糖含量,蛋白质和多糖含量的差异与菌体的生长特性和絮凝性相关。很多菌体EPS的多糖含量高于蛋白质含量,特别是在活性污泥的研究中,但有部分菌体的蛋白质含量高于多糖含量,如Acinetobactersp.YY-5[27]和Brevibacillussp.[26]等。王亮等[28]对1株白腐真菌(Phanerochaetechrysosporium)的EPS进行了研究,结果表明稳定期时的EPS为125 mg/L,其中蛋白质占30%,糖类占50%。Wang等[16]研究了1株异养反硝化菌(Enterobactersp.strain FL)的自聚集性能和胞外聚合物的关系,结果表明,该菌在12 h的自聚集率为17%左右,提取该菌的EPS发现,其蛋白质含量为每克干细胞中210 mg,多糖含量为每克干细胞中10 mg,其原因可能是该菌会产生较多的胞外酶。

表1 TN-10 EPS 中的蛋白质和多糖含量
注:以每克干细胞质量计。
2.4 胞外聚合物红外光谱图


图3 EPS的红外特征光谱Fig.3 FT-IR spectra of EPS
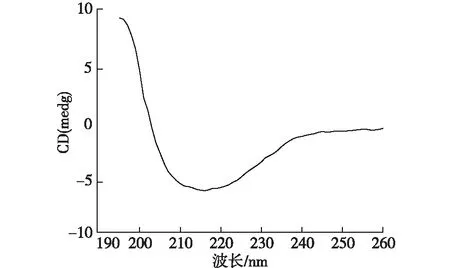
图4 EPS的圆二色谱图Fig.4 Circular dichroism spectra of EPS
2.5 圆二色谱测蛋白质二级结构
由于EPS中的蛋白质比多糖对絮凝性起着更重要的作用[16],而TN-10的EPS溶液中蛋白质含量高于多糖含量,使用圆二色谱对提取的胞外聚合物中蛋白质二级结构进行研究,结果如图4所示。由图4可知:在208和222 nm处的明显双峰是α-螺旋结构的特征峰,β-折叠在195和217 nm处有特征吸收峰,无规则卷曲在195 nm处有负的吸收峰[33]。利用CD pro程序包对圆二色谱的数据进行计算,其结果见表2。由表2可知:蛋白质中α-螺旋结构占主要比例,含量为55.08%。β-折叠和β-转角所占的比例分别为7.31和13.72%。菌体的絮凝性和自聚集能力与β-折叠、α-螺旋和三螺旋结构结构所占的比例成正相关,但是会受到无规则卷曲和β-反向折叠的抑制[34]。在TN-10的EPS中对絮凝性起促进作用的α-螺旋和β-折叠所占比例之和为62.39%,高于对絮凝性起抑制作用的无规则卷曲所占比例23.89%,这也是TN-10具有良好絮凝性的原因。
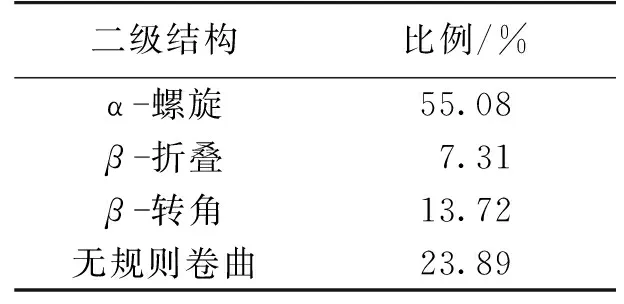
表2 EPS中蛋白质的二级结构
3 结论
笔者以1株硝化-反硝化菌Klebsiellasp.TN-10为研究对象,对其硝化-反硝化性能进行了考察,并对其絮凝性能进行了研究,主要研究结论如下。
2)研究了菌体的自聚集能力和聚集动力学,发现菌体在衰亡期的自聚集能力最强。
3)提取TN-10的胞外聚合物,测定其组成和含量,其中蛋白质含量为23.84 mg、多糖含量为18.64 mg(以每克干细胞质量计算)。
4)测定了EPS的红外光谱,并对EPS溶液中的蛋白质二级结构进行测定,其组成为α-螺旋55.08%、β-折叠7.31%、β-转角13.72%和无规则卷曲23.89%。
