邓恩桉开花结实期间内源激素含量变化
2019-02-13郭东强卢陆峰卢晨升尹明善蒙春江
郭东强,卢陆峰,卢晨升,尹明善,蒙春江
邓恩桉开花结实期间内源激素含量变化
郭东强1,卢陆峰2,卢晨升3,尹明善2,蒙春江2
(1.广西壮族自治区林业科学研究院/广西优良用材林资源培育重点实验室,广西南宁 530002;2. 环江毛南族自治县华山林场,广西环江 547100;3.玉林市林科所,广西玉林 537500)
植物的成花诱导、开花结实与内源激素的关系十分密切,单个激素的含量水平和激素间的动态平衡都对植株开花结实有着重要作用,研究内源激素变化情况,对于掌握植物开花结实机理和调控有着重要意义。为给邓恩桉开花结实研究与调控提供依据,采用酶联免疫法测定开花结实期间邓恩桉ABA、IAA、GA和ZR四种激素变化规律。结果表明:不同激素对于邓恩桉开花结实有着不同的影响,其变化规律也各异。低浓度的IAA有利于促进开花结实,GA对邓恩桉开花结实无显著影响,ZR和ABA的变化规律因个体差异而呈现不同规律,IAA/GA比值在促进邓恩桉开花方面起到重要作用。
邓恩桉;开花结实;内源激素
邓恩桉()是近年来得到广泛研究的耐寒桉树品种之一,是具有相当发展潜力的桉树品种。基于邓恩桉耐寒力强、生长速度快、材质优良等特性,国内开展了广泛的遗传育种研究[1-4]。
内源激素是植物体自身合成的产物,包括细胞分裂素、生长素、赤霉素、脱落酸和乙烯等5大类[5]。已有的研究认为,内源激素参与植物体内的多项调控作用,单个激素的含量和激素间的动态比值都有可能在植物生长发育的不同时期起到相应的重要作用,对植物的花芽分化和开花结实也有着重要作用[6-10]。鉴于国内邓恩桉开花结实过程中的内源激素含量变化的研究鲜有报道,本研究以在广西的邓恩桉种子园为对象,对其开花结实过程中内源激素含量和比值变化进行了分析,以期掌握邓恩桉开花结实规律和为开花结实生理基础提供理论依据。
1 材料与方法
1.1 材料
本研究选择邓恩桉嫁接种子园中往年开花结实量较多的3号、26号、29号、无性系单株作为对象,以不开花的28号单株作为对照。嫁接种子园建于2010年7月,位于广西环江县华山林场(25°18′N, 108°24′E),海拔800 m,为中山地貌,林地地势平缓,年平均气温16.2℃,极端最低气温-3.4℃,年降雨量1 475.6 mm,土壤为红壤,土层深60 ~ 80 cm。
1.2 采样方法
根据前期物候观察,邓恩桉在春季完成花芽分化,4-9月完成开花结实过程。本研究分别在2017年4月、6月、8月、9月开花结实期间采样,采集树体各方向上的正常生长叶片,洗净后将样品快速放入液氮罐中存储,带回实验室放置于-80℃超低温冰箱中保存待测。
1.3 内源激素测定方法
采用酶联免疫吸附测定法(ELISA)测定叶片中的生长素(IAA)、赤霉素(GA)、玉米素核苷(ZR)、脱落酸(ABA)含量。
1.4 数据分析
采用EXCEL 2017和SPSS 22.0进行数据整理、分析与作图。
2 结果与分析
2.1 邓恩桉开花结实期内源激素含量变化
2.1.1 IAA含量变化
由图1可知,IAA含量变化各异,不开花植株的IAA含量在开花结实期间先上升,在6月时达到最高值,为48.27 ng·g-1,而后呈下降趋势,花期结束后恢复至原水平。开花植株的IAA含量总体低于不开花植株,其中3号与29号呈现相同的“降-升-降”变化规律,而26号无性系则呈现缓慢增加的变化规律。6-8月是邓恩桉开花较为集中的时间,这一时间段内开花植株的IAA含量均呈上升趋势,而不开花植株则急剧下降。

表1 无性系间IAA含量变化规律 ng·g-1
2.1.2 GA含量变化
在GA的含量变化上,不开花植株的变化与开花植株截然不同。不开花植株的GA含量从4月开始增加,在6月达到峰值,而后到8月骤减,9月时含量上升。3号与26号GA含量变化规律相同,4-6月间呈下降趋势,而后变为上升趋势,8-9月间又变为下降趋势。29号无性系的GA含量在参试植株中的规律较为独特,在开花结实期间呈下降趋势,同时在开花结实期末期,各植株的GA含量在7.05 ~ 8.78 ng·g-1间,说明GA可能并不是影响邓恩桉开花结实的主要因子,且各无性系在营养生长阶段的含量无明显差异。

表2 无性系间GA含量变化规律 ng·g-1
2.1.3 ZR含量变化
ZR含量方面,开花植株中的3号与26号无性系的变化规律一致呈现增加的过程,而29号的ZR含量在4月时显著高于其他植株,达13.48 ng·g-1,并且呈现下降趋势,在开花结实末期才缓慢回升。不开花植株的ZR含量呈“升-降-升”的变化规律。

表3 无性系间ZR含量变化规律 ng·g-1
2.1.4 ABA含量变化
开花植株的ABA含量在开花结实初期在36.51 ~ 66.82 ng·g-1间,在6月时达到峰值,含量在73.53 ~ 99.82 ng·g-1间,而后呈先降后升的变化规律。不开花植株在开花结实初期时的ABA含量显著高于开花植株,为101.13 ng·g-1,则呈近乎于直线下降趋势,开花结实末期时与开花植株接近。
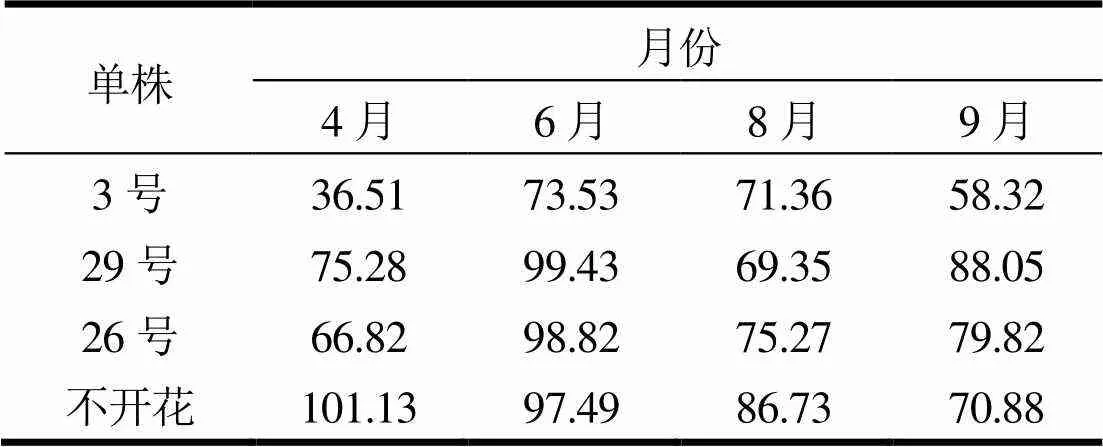
表4 无性系间ABA含量变化规律 ng·g-1
2.2 邓恩桉开花结实期内源激素比值变化
由图5可看出,不开花植株的IAA/GA值在6-9月间低于开花植株,说明IAA/GA比值在促进邓恩桉开花方面起到重要作用;ABA/GA值的变化上,不开花植株和3号植株的变化曲线几乎一致,而26号和29号的变化趋势相反,可推测ABA/GA在邓恩桉开花结实方面不起关键作用;IAA/ZR值与ABA/ZR值的变化无明显规律性。
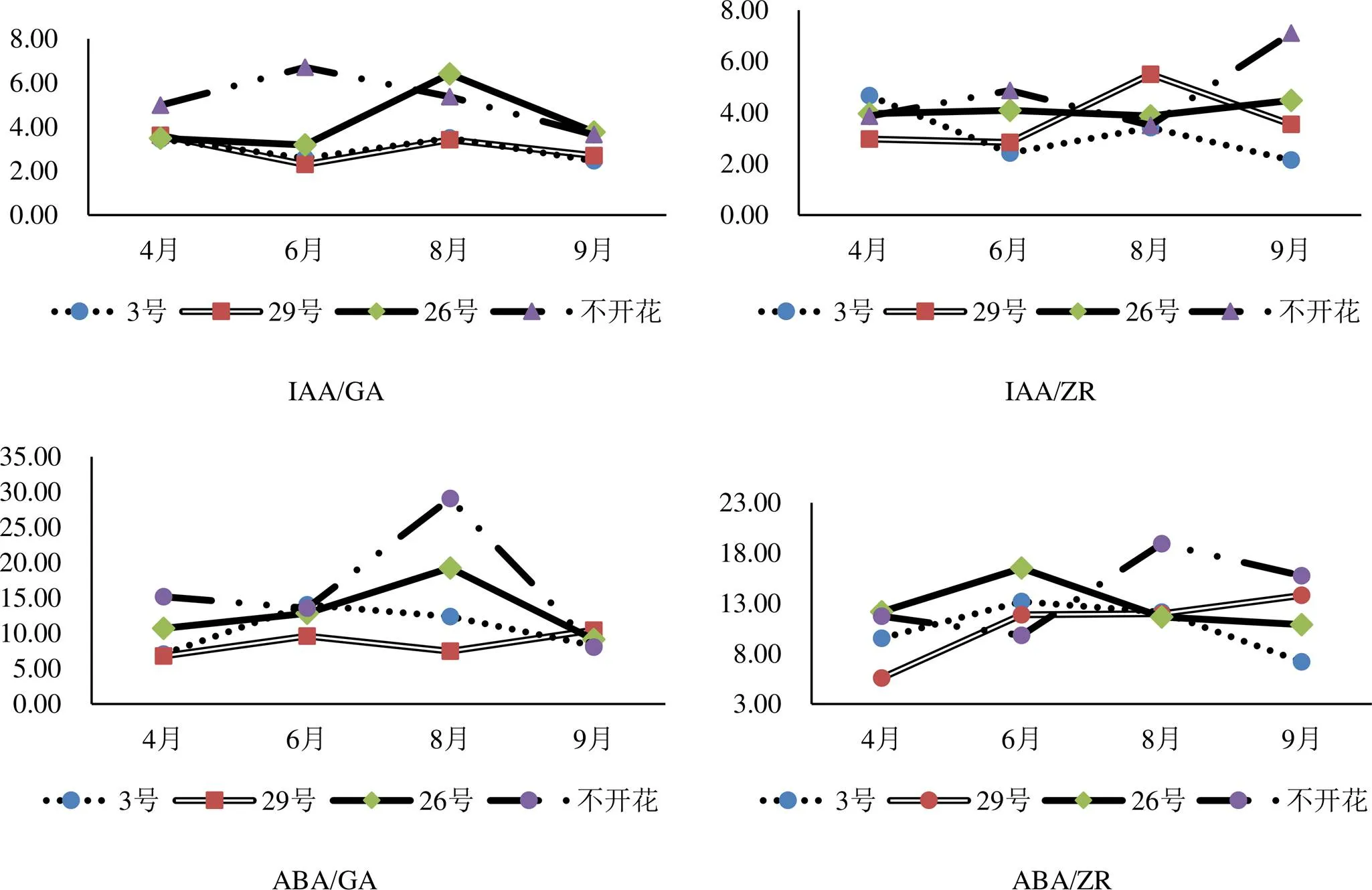
图5 邓恩桉开花结实期间不同内源激素比值变化趋势
3 结论与讨论
3.1 内源激素含量变化与邓恩桉开花结实的关系
植物的花芽分化和开花结实,是营养生长转变到生殖生长的结果,这一转变过程是植物生长到一定阶段,在适宜的外部条件下,通过复杂的形态结构和生理变化而实现,其中植物内源激素发挥了重要的调控作用,且这种调控作用并不是由某一激素单独作用而产生的[11-13]。
IAA为植物生长内源激素的一种,既具有促进植物生长的作用,也具有抑制植物生长、促进器官建成的作用。关于IAA在植物开花结实过程中的作用,目前并未有一致的研究结论,部分学者认为,IAA 是花芽形成的抑制因子之一,但也有学者发现较高浓度的IAA含量对植物花芽的分化有促进作用[14-16]。对杏花、茉莉花和金茶花的研究结果表明,高浓度IAA有助于蕾的生长,低浓度IAA可促进花的开放和衰老[17-19]。本研究发现,邓恩桉开花植株的IAA含量总体和变化幅度均低于不开花植株,说明低浓度的IAA对其开花结实有促进作用。
赤霉素对种子萌发和植物生长均有促进作用,而其对于开花结实调控的作用因植物而异[20-21]。GA对于植株的花芽分化和开花结实具有抑制作用,并且在花期时促进花的凋谢和脱落。本研究中GA在开花与不开花植株间、开花无性系间的变化规律各异,说明GA并不是邓恩桉开花结实的主导因子。
玉米素核苷是细胞分裂素(CK)中一个较活跃的种类[22],多数研究表明ZR对于开花结实有促进作用,同时能够促进植物细胞分裂和增大。本研究中,开花和不开花植株在花芽分化过程中的ZR积累程度不一,不开花植株在开花结实期内的ZR含量呈“升-降-升”的变化趋势,而开花植株因无性系不同而有所区别。
通常认为,脱落酸最主要的生理作用是促进器官脱落和衰老并抑制植株生长。关于ABA 对开花的作用,部分学者认为随着开花过程的进行,ABA含量持续增加,直至花、果等器官凋落,秦建彬等[23]、OKUDA[24]等在研究中发现,ABA的升高利于开花;ABA 可以同GA3产生拮抗作用,促进花芽孕育。邓恩桉开花植株中ABA含量变化呈现两极差异,3号和26号无性系花芽分化期积累的ABA显著低于29号,且3号与26号无性系ABA含量随着开花结实进程逐渐增加,29号无性系则逐渐减少。邓恩桉开花植株个体间的差异,导致ABA含量呈现不同变化规律。
3.2 内源激素平衡与邓恩桉开花的关系
植物内源激素含量对植物开花结实起着关键作用,研究表明单一激素并不能完全决定植株开花结实,各激素间的动态平衡才能诱导植株花芽分化和开花结实[25]。本研究结果表明,IAA/GA比值在促进邓恩桉开花方面起到重要作用。
综上所述,邓恩桉开花结实受到单个激素变化和激素间动态平衡的影响,开花植株与不开花植株间、开花无性系间均存在不同程度的差异。在施用外源激素促进邓恩桉开花结实时,需要根据单株差异有针对性地进行,才能收到较好的效果。
[1] 郭东强,叶露,周维,等.2个种源邓恩桉木材纤维特性及变异[J].浙江农林大学学报,2014,31(4):502-507.
[2] 张建明.广西桂林耐寒桉树引种试验研究[J].桉树科技,1996(1):44-50.
[3] 项东云,陈健波,陈崇征,等.广西耐寒桉树研究进展[J]. 广西林业科学,2002,31(4):178-180.
[4] 陈健波,张照远,项东云,等.邓恩桉优树的选择标准[J].林业科技开发,2008,22(1):17-20.
[5] 王丽萍,李志刚,谭乐和,等.植物内源激素研究进展[J].安徽农业科学,2011,39(4):1912-1914.
[6] 马亮,李建安,刘丽娜,等.油桐开花期内源激素含量的变化[J].经济林研究,2009,27(2):31-34.
[7] 艾星梅,胡睿宇,胡燕芳.马铃薯开花与内源激素动态变化的关系[J].西北植物学报,2018,38(1):87-94.
[8] 朱振家,姜成英,史艳虎,等.油橄榄成花诱导与开花期间侧芽激素含量变化[J].林业科学,2015,51(11):32-39.
[9] 何见,蒋丽娟,李昌珠,等.光皮树开花过程中内源激素含量变化的研究[J].中国野生植物资源,2009,28(2):41-45.
[10] 孔德政,靳丹丹,何松林,等.碗莲开花的解剖学以及内源激素变化规律的研究[J].北京林业大学学报,2009,31(3):42-45.
[11] RAKNGAN J, GEMMA H, IWAHORI S. Flower bud formation in Japanese pear trees under adverse conditions and effects of some growth regulators [J]. Journal of Japanese Tropical Agriculture, 1995, 39(1):1-6.
[12] ZHANG D, REN L, YUE J H, et al. GA4and IAA were involved in the morphogenesis and development of flowers inpraecox ssp orientalis[J]. Journal of Plant Physiology,2014,171(11):966-976.
[13] 崔澂,吕忠恕,陆嘉陵.第十二届国际植物生长物质会议简况[J].植物生理学通讯,1986,1(2):66-70.
[14] 吴邦良,夏春森,赵宗方.果树开花结实生理和调控技术[M].上海:上海科学技术出版社,1994.
[15] 黄卫东,原永兵,彭宜.温带果树结实生理[M].北京:北京农业大学出版社,1994.
[16] 王海波,赵君全,王孝娣,等.新梢内源激素变化对设施葡萄花芽孕育的影响[J].中国农业科学,2014,47(23):4695-4705.
[17] 杨途熙,王佳,魏安智,等.杏的开花生理研究[J].西南大学学报(自然科学版),2010,32(12):25-31.
[18] 高丽萍,夏涛,张玉琼,等.茉莉花发育及开放期间内源激素研究[J].茶叶科学,2002,22(2):156-159.
[19] 漆小雪,韦霄,王熊军,等.金花茶花期内源激素含量的变化[J].江苏农业科学,2013,41(3):141-144.
[20] 黄羌维.龙眼内源激素变化和花芽分化及大小年结果的关系[J].热带亚热带植物学报,1996,1(2):58-62.
[21] 王玉华,范崇辉,沈向,等.大樱桃花芽分化期内源激素含量的变化[J].西北农业学报,2002,1(1):64-67.
[22] 傅永福,孟繁静.植物的成花生理信号(综述)[J].中国农业大学学报,1998,3(3):1-11.
[23] 秦建彬,魏翠华,余祖云,等.大花蕙兰开花与激素关系的研究[J].中国农学通报,2011,27(31):109-112.
[24] OKUDA H, KIHARA T, IWAGAKI I. Effects of cropping on photosynthesis, dark respiration, leaf ABA concentration and inflorescence induction in[J]. Engei Gakkai Zasshi,1995,7(64):9-16.
[25] 宗学凤,李芳红,张建奎,等.蓝粒小麦植株发育进程中内源激素的变化[J].西南大学学报(自然科学版),2007,29(12):58-62.
Endogenous Hormone Variation induring Flowering and Fruiting
GUO Dongqiong1, LU Lufeng2, LU Chensheng3, YIN Mingshan2, MENG Chunjiang2
(; 2; 3.)
The induction of flower formation, flowering and then fruit production by plants is sensitively affected by endogenous hormones. The concentration of a single hormone and the dynamic balance between hormones can have important effects on flowering and subsequent fruiting. Studying temporal changes in endogenous hormones and grasping the mechanism of flowering and fruiting in plants along with the regulation of this process is of great importance. In order to facilitate research into and the regulation offlowering and fruiting, an enzyme-linked immunosorbent assay was used to determine the regulation of four hormones ABA, IAA, GA and ZR in this species during flowering and fruiting. The results showed that different hormones had different effects on the flowering and fruiting of, and the patterns in the changes of concentration of each hormone during the process also varied. Low concentration of IAA is beneficial to promote flowering and fruiting, whilst GA seemed to have no significant effect on flowering and fruiting of. The change of ZR and ABA shows different rules due to individual differences, and the ratio of IAA/GA seems to play an important role in promoting flowering in.
;flowering and fruiting; endogenous hormones
Q945
A
10.13987/j.cnki.askj.2019.04.004
广西优良用材林资源培育重点实验室课题(15-A-01-01)
郭东强(1986— ),男,硕士,工程师,主要从事桉树遗传育种与栽培,E-mail:79244381@qq.com
