甜瓜蛋白激酶类基因CmPKC的克隆、表达及遗传转化分析
2019-01-18叶春秀赵增强张国丽庄振刚谢宗铭
叶春秀,赵增强,张国丽,于 航,庄振刚,谢宗铭*
(1.新疆农垦科学院生物技术所/作物种质创新与基因资源利用兵团重点实验室,新疆 石河子 832000;2.新疆农业大学,新疆 乌鲁木齐 830000)
【研究意义】蛋白质的可逆磷酸化是生物体中普遍存在的一种调节机制,它几乎涉及所有的生理和病理过程。如光合作用、基因表达、细胞的生长发育、糖代谢、神经递质的合成与释放、甚至癌变等等。据估计,真核生物1 %~3 %的基因编码蛋白激酶,而植物中可能有2 %~3 %的基因编码蛋白激酶,这些蛋白激酶参与了环境刺激和植物激素等多种信号在细胞内的传递[1-3]。虽然植物蛋白激酶的研究发展迅,但仍然远远落后于动物和人类,许多蛋白激酶的生物学功能还不清楚,但也有所进展,近来在植物中也发现了分裂原激活的蛋白激酶(Mitogen-actived Protein Kinase, MAPK)的相似蛋白,MAPK(ERKs)是一族胞浆内广泛分布的含有丝氨酸/苏氨酸残基的蛋白质激酶[4-9]。MAPK 受双重磷酸化,是真核蛋白激酶的一种特殊分类,能在丝氨酸、苏氨酶、酪氨酸残基上磷酸化。在MAPKs 的活化环中激活的磷酸化发生在保守的Thy-X-Tyr 基元中。MAPK 通过使转录因子磷酸化而改变基因的表达水平。MAPK 是真核信号传递中的关键因素,MAPKs 可能参与各种发育或胁迫信号转导的磷酸化级联反应。有研究表明:一些植物生长因子如植物生长素、乙烯可以活化MAPK等。另外,研究表明小麦上TaLRK蛋白激酶家族基因能够参与小麦抗白粉病反应[3]。【前人研究进展】甜瓜作为新疆的特色水果之一,其蛋白激酶的研究将为甜瓜的基础代谢、抗逆和抗病虫害的机理研究奠定基础,并且目前甜瓜蛋白激酶的研究还属于零星研究,远未能构成完整的反应网络[10]。【本研究切入点】本项目组根据NCBI上已报道的其它物种的蛋白激酶类基因,比对相对保守的序列,设计相关的引物,采用RT-PCR方法获得了CmPKC的部分序列,根据部分序列的测序结果设计引物,分别采用3’ RACE和5’ RACE技术获得了该基因的全长序列。【拟解决的关键问题】该基因的克隆与表达分析将为进一步研究蛋白激酶类基因在抗病性调控过程中所起的作用及下一步构建植物表达载体及基因沉默奠定研究基础。
1 材料与方法
1.1 材料与试剂
抗白粉病与不抗白粉病的甜瓜材料由新疆兵团第六师科学研究所提供,种植于新疆农垦科学院生物技术研究所实验室。
RNA Trizol提取试剂盒、TaqDNA 聚合酶、DNA 回收纯化试剂盒、质粒提取试剂盒、蛋白质分子量标记、T4DNA连接酶购自天根公司;3’及5’-RACE试剂盒、克隆载体 pMD 19-T Vector购自 TaKaRa 公司;荧光定量试剂盒购自全士金公司;Thermo Scientific RevertAid First Strand cDNA Synthesis Kit购自Thermo公司。
1.2 试验方法
1.2.1 总RNA的提取 总RNA的提取采用天根Trizol试剂盒,根据操作说明完成。材料为2对真叶期的白粉病抗病与不抗病的甜瓜幼苗及接菌0,1,5,9,24 h不同抗性的材料,液氮速冻,-80 ℃保存用于RNA的提取。
1.2.2 cDNA的合成及基因部分序列的克隆 参照Thermo Scientific RevertAid First Strand cDNA Synthesis Kit 合成cDNA第一链,根据相对保守序列设计引物,进行部分序列的扩增。
1.2.3 3’ RACE和5’ RACE 参照TARAKA试剂盒说明书,以Total RNA为模板,使用3′ RACE adaptor引物进行反转录反应合成lstStrand cDNA,使用上游外侧特异性引物(GSP1,表3)和3′ RACE Outer Primer进行1stPCR反应,再使用上游内侧特异性引物(GSP2)和3′ RACE Inner Primer进行2ndPCR反应。PCR产物连接PMD9-T进行TA克隆.挑选阳性克隆,送给公司测序。
5’RACE。使用Alkaline Phosphatas去掉Total RNA 中裸露的5’磷酸基团. 用Tobacco Acid Pyrophosphatase去掉mRNA 的5’帽子结构,保留一个磷酸基团。使用T4RNALigase将5’RACE Adaptor连接到去帽后的mRNA 上,然后以此mRNA 为模板,使用Reverse Transcrip-tase M-MLV (RNase H.),Random 9 mers进行反转录反应合成cDNA。使用下游外侧特异性引物GSP3和5’ RACE Outer Primer进行Outer PCR反应,再使用下游内侧特异性引物GSP4和5′RACE Inner Primer进行Inner PCR反应 PCR产物连接PMD19-T,TA克隆,挑选阳性克隆,送公司测序。
1.2.4 荧光定量及半定量检测 利用实时荧光定量 PCR 的方法检测CmPKC基因在不同抗性甜瓜材料及不同接种时间点的相对表达量。上游引物序列为:5’-CTTGCCTTCTTTGCTTGG-3’;下游引物序列为:5’-TCTTCTTGGCGGAGTGGT-3’。用PR0961402 36实时荧光PCR 检测系统进行检测,反应体系如下:10 μl 2× SYBR®Premix ExTaqTM,正反向引物均为 0.4 μl,cDNA 模板 2 μl;加 ddH2O至 20 μl。反应程序:95 ℃ 3 min;95 ℃ 10 s,58 ℃ 20 s,72 ℃ 30 s,35 个循环,实时 PCR反应以甜瓜18S rDNA为内参,每个反应重复 3 次。
1.2.5 烟草转化及验证 采用常规的叶盘法及农杆菌转化方法进行烟草的转化,获得转化烟草植株后进行阳性检测,检测引物采用部分基因序列引物和载体NPTII引物进行,将同时扩增出条带的植株定位阳性植株,进行移栽,收获种子保存,用于后期实验。
2 结果与分析
2.1 甜瓜RNA的提取
本实验根据甜瓜叶片特性采用天根Trizol试剂盒进行RNA的提取,参照说明书进行相应的操作,采用1 %的琼脂糖凝胶进行RNA质量与浓度的检测,检测结果表明提取的RNA浓度与质量较好,能够满足后期的实验需要(图1)。
图1 RNA提取质量检测Fig.1 Quality of RNA of Cucumis melon L.
2.2 甜瓜CmPKC基因中间片段的扩增
以不同抗性甜瓜材料的cDNA为模板,以中间部分序列引物进行扩增,扩增产物经电泳检测发现在约2600 bp处有一条亮带(图2)。将此片段进行回收纯化,连接PMD19-T Vector 连接,转化检测后进行测序,测序结果显示该片段的大小为2500 bp,与其他植物的类似蛋白激酶类基因同源性较高。
2.3 甜瓜CmPKC基因3’-RACE和5’-RACE
通过3’RACE和5’RA CE分别获得了约500bp的3’端片段和400 bp的5’端片段(图3)。测序结果表明,其序列前后分别能与CmPKC基因中间序列前后吻合,说明获得了甜瓜CmPKC基因的3’端和5’端,然后将中间片段,3’端和5’端片段拼接,设计全长引物进行扩增,最终获得的CmPKC基因长约为2914 bp,开放阅读框为2493 bp,编码831个氨基酸(图3)。

图2 CmPXC基因中间部分序列扩增图谱Fig.2 PCR amplification of CmPKC of Cucumis melon L.
2.4 CmPKC蛋白的氨基酸序列分析、比对与进化树的构建
如图4所示,利用NCBI网站数据库对CmPKC的同源基因氨基酸序列进行比对和搜索,结果显示,CmPKC与已报道的香瓜蛋白激酶不同剪接体,黄瓜、杨树、欧洲油菜、羽衣甘蓝、芜菁等植物的蛋白激酶类似基因氨基酸具有较高的同源性,其中与甜瓜G-type lectin S-receptor-like serine/threonine-protein kinase 剪接体2同源性最高,达到88 %。
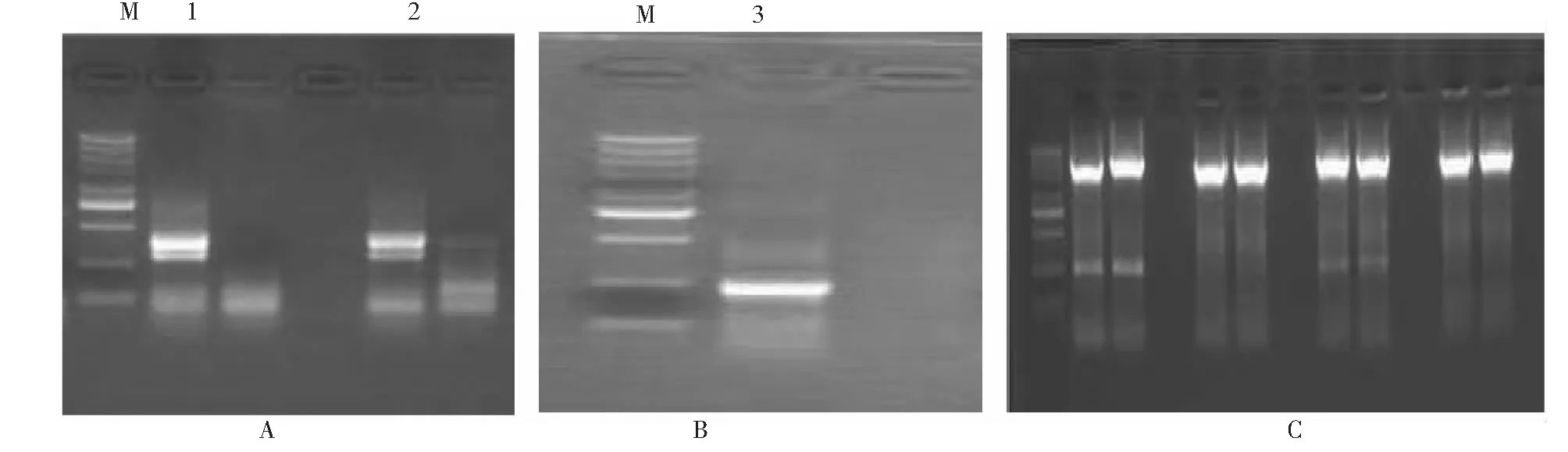
A.3′-RACE;B.3′-RACE;C.Full length图3 甜瓜CmPKC基因3’ RACE,5’ RACE和全长序列扩增结果Fig.3 PCR amplification of 3’, 5’end and full length of CmPKC gene
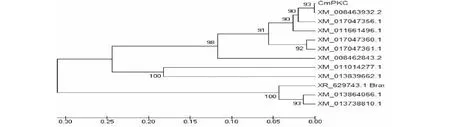
The amino acid sequences used to build up the phylogenic tree were: Cucumis melo transcript variant X2(XM008463932.2); Cucumis melo transcript variant X1(XM017047356.1); Cucumis melo transcript variant X3(XM017047360.1); Cucumis melo transcript variant X4(XM017047361.1); Cucumis sativus mRNA (XM011661496.1); Cucumis melo mRNA (XM008462843.2); Populus euphratica(XM011014277.1); Brassica napus RKS1(XM013839662.1); Brassica napus SD2-5(XM013864066.1); Brassica pleracea SD2-5(XM013738810.1); Brassica rapa SD2-5(XR629743.1)图4 CmPXC与同源基因编码蛋白的系统进化树Fig.4 Phylogenic tree of CmPKC and protein coded by by its homologous genes
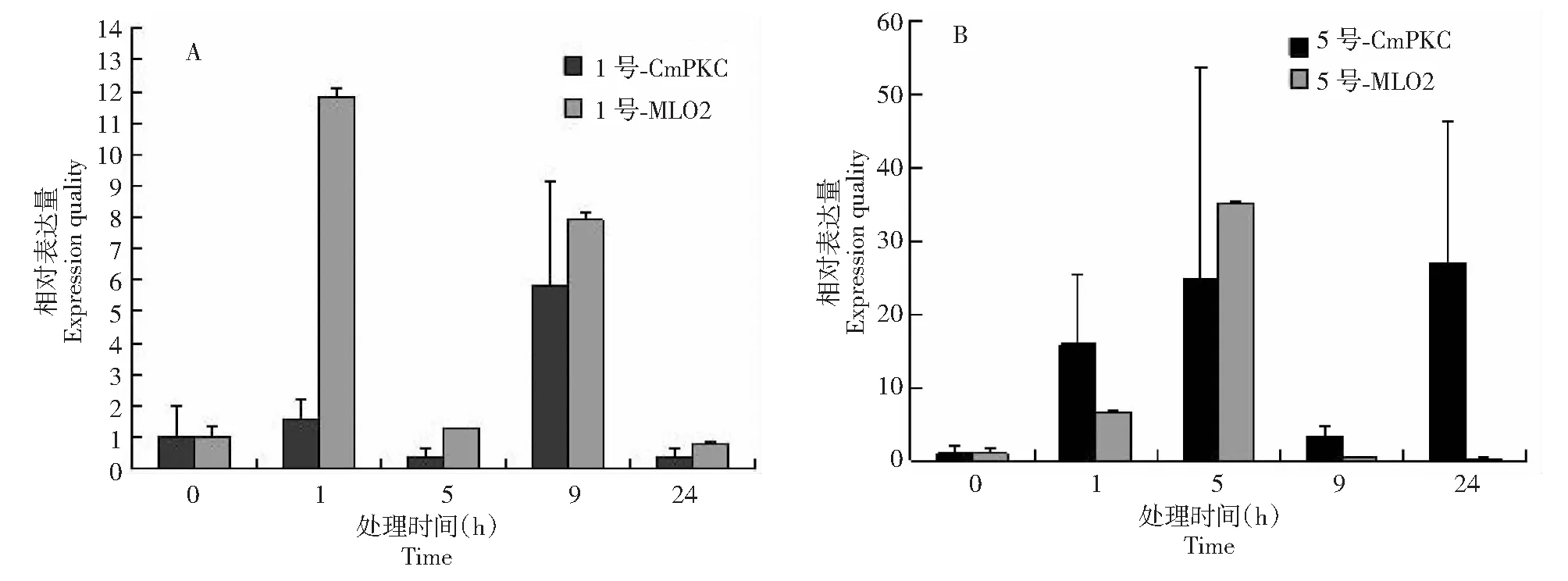
A:抗性材料接菌后相对表达量;B:感病材料接菌后相对表达量A:Resistance material;B: Susceptibility material图5 CmPKC基因在甜瓜白粉病抗性及感病材料上接菌后的相对表达量Fig.5 Expression quality of CmPKC in different material of Cucumis melon L.
2.5 甜瓜CmPKC基因在不同抗性材料上的表达分析
利用甜瓜的18S作为内参,对CmPKC基因在不同抗性材料上进行荧光定量PCR分析,结果表明在抗性材料上,接菌9 h后该基因的表达量最高(图5A),而在感病材料上接菌5 h后该基因的表达量最高(图5B),与现有报道的一个与甜瓜感病相关MLO家族基因表达量变化规律一致,但表达量远远不及该家族基因,说明所获得的基因可能与甜瓜白粉病的感病相关,且属于早期应答基因类型;通过对不同接菌时间的抗病与感病材料CmPKC基因转录水平定量分析显示该基因在甜瓜白粉病上的抗性机制类似于MLO基因在植物上对白粉病的抗性机制,在抗性材料上在接菌24 h前出现的表达高峰可能与乳突反应的发生有一定的联系,当完成乳突反应后,病原菌即被阻隔在细胞之外直至失去侵染活性,随即表达量下降。而感病品种的表达量在24 h 后开始上升,且高峰出现在72 h。由于24 h 内是白粉菌完成侵入的关键时期,在抗病品种24 h前出现了表达高峰,推测抗病材料中CmPKC基因可能直接参与了抗病反应。而感病品种由于峰值反应的延迟,错过了抵抗病原菌侵入的最佳时机,导致了植物感病(图6)。
2.6 烟草的遗传转化
提取转化烟草及野生烟草植株的基因组DNA,采用基因部分序列引物及NPTII基因引物分别进行PCR检测,最终选择能够同时扩增出目的基因和NPTII基因的植株定位阳性植株(图7),经过后期观察,该基因在起抗逆作用同时还具有促进烟草开花的功能,其较野生植株开花期提前很多(图8)且抗逆性有所增加,其在抗病过程中所起的作用有待深入分析和研究。

图7 转基因烟草植株目的基因及NPTII鉴定Fig.7 PCR amplification of CmPXC and NPTII

图8 转基因烟草提前开花植株与对照植株Fig.8 Comparation of standard and early flowering plants
3 讨 论
在植物体内,存在着大量参与植物组织形成和器官发育的激酶。 拟南芥中的ERECTA基因编码含LRR 的激酶,其与器官的形成及营养生长有关[10];HAESA也是一种含LRR 的激酶,参与了花组织器官的脱落[11];CLAVATA与根和花的分生组织中细胞的增殖和组织形成有密切的关系[12];另外,WAK(wall2associated receptor2like kinase) 通过细胞的伸长生长,调控侧根的生长发育[13];AtSERK1编码一种参与体细胞胚胎发生的类受体蛋白激酶;SCM和根的表皮细胞发育有关[14-15]。矮牵牛中的PRK1在花粉粒中特异表达,在小孢子减数分裂后的花粉发育和授粉中起信号传递作用,保证花粉粒正常发育和萌发[16];玉米的Crinkly4 基因编码的激酶参与调控叶表皮细胞和胚乳中糊粉细胞的分化[17]。
另外,已鉴定的抗病基因中有多个类受体激酶,对该类抗病基因的克隆及其序列分析和功能的研究,促进了人们对激酶在植物抗病中作用的认识。水稻抗白叶枯病基因Xa21 和Xa26都编码一个具有胞外LRR 区的丝/苏氨酸激酶,LRR 区参与蛋白质相互作用[18-19]。水稻抗稻瘟病基因Pi2d2编码的蛋白质胞外是B 凝集素(B2lectin) 结构,胞内是丝/苏氨酸激酶区[20]。Martin 等从番茄中克隆的抗丁香假单孢杆菌(Pseudomonassyingaepv.tomato) 的基因Pto,编码一个细胞内的蛋白激酶[21]。 拟南芥的FLS2 基因,编码含LRR 结构域的类受体激酶,在植物抗病和病原菌识别中起重要作用。大麦的Rpg1基因编码的激酶抗锈病[22]。小麦抗叶锈病基因Lrk10 编码胞外含有S结构域的受体蛋白激酶[23-24]。
本文通过同源基因克隆,步移和RACE法成功克隆出CmPKC基因全长,其开放阅读框为2493 bp,编码831个氨基酸,与已报道的甜瓜蛋白激酶不同剪接体,黄瓜、杨树、欧洲油菜、羽衣甘蓝、芜菁等植物[8]的蛋白激酶类似基因氨基酸具有较高的同源性,其中与甜瓜G-type lectin S-receptor-like serine/threonine-protein kinase 剪接体2同源性最高,达到93 %。对不同抗性的甜瓜材料进行白粉菌接种后,半定量和荧光定量分析结果显示该基因与目前报道的甜瓜MLO基因家族表达量在不同抗性材料上的变化趋势一致,推测该基因在甜瓜白粉病上的抗性机制可能类似于MLO基因在植物上对白粉病的抗性机制,且属于早期应答基因,即在抗性材料上在接菌24 h前出现的表达高峰可能与峰值反应的发生有一定的联系,当完成峰值反应后,在表面产生凸起,病原菌即被阻隔在细胞之外直至失去侵染活性,随即表达量下降。而感病品种的表达量在24 h 后开始上升,且峰值出现在72 h。由于植物24 h内是白粉菌完成侵入的关键时期,在抗病品种24 h前就出现了表达高峰,推测抗病材料中的CmPKC基因可能直接参与了抗病反应,而感病品种由于峰值反应的延迟,错过了抵抗病原菌侵入的最佳时机,导致了甜瓜感病,具体调控途径有待于进一步分析。转基因烟草鉴定显示该基因可能还参与植株的生长发育过程,还需后期经过进一步的鉴定确定其具体参与的调控路径。
