油菜及其近缘植物形态标记性状基因定位与克隆研究进展
2019-01-17刘忠松
杨 柳,康 雷,刘忠松
(湖南农业大学农学院,长沙 410128)
油菜主要用作油料作物,近年来也作为观赏作物、蔬菜作物、饲料作物等种植。从植物学角度看,油菜包括白菜型油菜、芥菜型油菜和甘蓝型油菜三类。三类油菜在形态上存在明显差别,如白菜型油菜较矮;甘蓝型油菜、芥菜型油菜较高;白菜型油菜、芥菜型油菜有天然的紫叶、多室、黄籽特征等。这些性状差异特异性强、易识别、受环境条件影响小,不仅可以作为标记性状直接应用于遗传研究、杂种鉴别、种子纯度鉴定等诸多方面,而且有些还与作物的产量、品质甚至抗逆性相关联。近年形态标记性状作为重要的育种选择参考越来越受到重视,如随着观赏油菜的兴起,对油菜品种的叶型、叶色、株高和花色都提出了新的要求[1,2]。
油菜的根、茎、叶、花、果、实的形态性状研究由来已久,但以往研究大多停留在对表观性状遗传分析方面。近年随着油菜及近缘植物全基因组测序完成和序列公布,已将控制叶色、叶型、株高、花色、种子颜色等性状的基因进行了定位和克隆,从而为比较不同油菜类型间控制同一性状的基因差异提供了条件。本文仅对油菜及其近缘种的形态标记性状基因定位和克隆进展进行评述。
1 叶色
油菜叶片一般为绿色,但有紫色、黄色、桔黄色等变异类型。紫叶白菜含有更多的芦丁[3],桔黄叶白菜积累更多的β-胡萝卜素[4],这些物质提高了白菜的营养价值。在油菜(Brassica napus)、芥菜(B.juncea)、白菜(B.rapa)和甘蓝(B.oleracea)等芸薹属植物中已经发现了有天然的紫叶突变,属于单基因显性突变,并且紫叶性状可通过种间杂交由芥菜、白菜转移到油菜和甘蓝中去[5,6]。油菜控制紫叶性状的基因在A03染色体,编码APR2基因[7],芥菜控制紫叶性状的基因位于B02染色体(重新命名为B05[8]),编码MYB转录因子基因[9],白菜控制紫叶性状的基因定位到A03、A07和A09染色体[10],A03染色体的编码基因为LBD39[10],A09染色体有2个控制叶色的基因,分别为DFR和bHLH类转录因子EGL3[11,12],甘蓝控制紫叶性状的基因定位到C03、C06和C09染色体,C06染色体上的紫叶基因为MYB2和MYBL2,而C09染色体的紫叶基因为DFR[13~19]。紫叶的形成是由于过量积累花色素[20~23]。花色素属于类黄酮类物质。类黄酮的生物合成途径研究成果颇丰,在芸薹属植物中有数十个基因参与这一途径[23,24],还有许多基因没有找到自然突变体。
桔黄叶是大白菜的重要性状,多个研究都把单基因隐性遗传的桔黄叶指向A09染色体的影响胡萝卜素合成的CRTISO基因[25~29]。芸薹属植物像水稻等植物一样,有许多叶片黄化突变体,在叶绿体基因组[30]和C01、C07和C08染色体定位有黄化基因[31~34],但至今仅1个油菜黄化突变体和1个白菜黄化突变体基因被克隆,分别编码血红素加氧酶1(PHO1)基因和叶绿体rps4基因。白菜QTL定位及基因注释分析认为,SCO3、ABI3、FLU、HCF153、HEMB1和CAB3等基因与叶色和叶绿素合成有关[35]。
2 光叶
油菜叶片表面一般会合成、积累一层蜡粉,保护植株免受干旱、高温等环境胁迫。蜡质是链非常长的脂肪酸(C20~C34)衍生物质,在油菜叶片中检出有24种之多(C24~C31)[36]。如果叶片表皮细胞不合成蜡质,叶片表面就因不能积累蜡粉而变成光叶(glossy)。光叶白菜、甘蓝口感更好,而且不利于蚜虫等多种昆虫取食[37]。光叶既有显性遗传的,也有隐性遗传的。油菜中单基因显性光叶性状是由A09染色体CER1基因单碱基突变所致[38],白菜中克隆了A01和A09染色体上CER2和CER1基因[39,40],甘蓝中克隆了C01和C08染色体上的CER4、CER1基因,其中CER1基因有3个等位突变[41~45],而C03染色体有涉及蜡质合成的O-酰基转移酶家族蛋白(WSD1-like)、3-酮酰CoA合成酶(KCS)和脂肪酰CoA还原酶(FAR)3个候选基因[37]。拟南芥中涉及蜡质合成、分泌和调控的基因有20余个[46],通过生物信息手段分析其中10个基因的油菜同源基因,发现有44个拷贝,除了CER10外,每一个拟南芥基因在油菜中有4个或更多同源基因[47]。
3 叶型
花叶是由于叶片深度缺刻造成,在油菜、芥菜中均有花叶自然突变体。油菜花叶是单基因不完全显性遗传,由于A10染色体LMI1基因启动子区域插入突变所致[48,49]。白菜QTL定位及基因注释分析认为CYCD3;1、CYCB2;4、AN3、ULT1和ANT基因与叶片大小有关[35]。遗传基因组学研究发现,白菜A03、A09和A10染色体分别携带有控制叶型的KRP2、ERECTA和LONGIFOLIA1等基因[50]。
4 矮秆
甘蓝型冬油菜株高一般在180~200 cm,容易倒伏,不利于机械收获。由于油菜收获指数低(0.28~0.35),所以一些育种家认为油菜植株应高大。但矮秆是水稻、小麦等作物第一次“绿色革命”的目标性状,油菜能否培育出矮秆高产品种近年受到重视。控制植物矮秆性状的基因涉及到赤霉素、生长素、油菜素内酯和独脚金内酯的合成、转运和传导。油菜中现有矮秆突变体多是化学诱变产生的碱基替换的结果,株高在70~100 cm,表现为单基因半显性遗传。已经定位的矮秆基因位于油菜A03、A06、A09和C05、C07、C09染色体上,A03染色体的矮秆基因编码控制生长素合成的IAA7[51],C05染色体的矮秆基因编码控制赤霉素合成的GID1[52],其他已克隆的油菜、白菜矮秆基因都编码控制赤霉素传导的RGA基因[53~59]。油菜是无限花序作物,如果变成有限花序,株高也会降低。决定油菜有限花序的基因定位在A10染色体,编码TFL1基因[60,61]。通过株高QTL定位、整合和候选基因鉴定,发现油菜有287个控制株高的候选基因[62]。
5 花色
油菜种植面积大、花期长、花色鲜艳,近年来以观赏油菜花为主题的油菜花节助推了乡村旅游的兴起,需要培育红色、紫色、彩色等不同花色的油菜品种。传统的油菜花一般是黄色(鲜黄色、金黄色、桔黄色),通过远缘杂交已将甘蓝(白花芥蓝)、萝卜、诸葛菜等植物的白花、紫花和红花等性状导入到油菜中。影响花色的色素物质主要有三类,即蓝色到红色的花青素、红色的甜菜素和黄色到带红色的类胡萝卜素,这些色素的组成和含量决定花色的变化[63]。油菜桔黄色花是由2个隐性基因位点控制,其中一个位点定位在C09染色体,编码类胡萝卜素代谢途径玉米黄素环氧化酶ZEP基因[64]。由白花芥蓝转移到油菜的白花性状是单基因显性遗传,基因定位在C03染色体,编码类胡萝卜素裂解双加氧酶基因CCD4[65,66],白花是由于转座子插入所致。芥菜乳白花性状是由2个隐性基因控制,分别定位在A02和B04(B06[8])染色体,编码硫酯酶家族蛋白PES2和PES1基因[67,68]。
6 种子颜色
油菜未成熟种子为绿色,成熟种子为黑色、褐色或黄色。在相同遗传背景下,油菜黄色种子比黑色、褐色种子含油量更高,种皮更薄,不含原花色素。因此选育黄籽油菜品种是重要的育种目标。在自然界还没有发现甘蓝型黄籽油菜,将白菜型油菜、芥菜型油菜、白芥(Sinapis alba)中发现的黄籽性状通过种、属间杂交转移到油菜中,培育出了甘蓝型油菜黄籽类型。白菜和白菜型油菜黄籽性状基因分别定位在A06和A09染色体,A06染色体TTG1基因调控种子颜色[69,70],A09染色体有2个编码转录因子的基因TT1和TT8被认为调控种子颜色[71~73]。芥菜型油菜黄籽性状基因定位在A09和B03(B08[8])染色体[74,75],同样编码TT8基因,但芥菜型油菜黄籽很可能不是来源于白菜型油菜黄籽,因为突变位点不同[71,75]。
7 多室
油菜一般由1个假隔膜将角果分为2室,但也发生了天然的3或4室突变,这种具有3或4室的油菜称为多室油菜。多室油菜每角果种子数增加,有利于提高单产。白菜型黄籽油菜中突变形成的多室油菜是由于A04染色体CLV3基因单碱基替换所致[76,77],而芥菜型黄籽油菜中突变形成的多室油菜是由2个隐性基因控制,分别位于A07和B07(B03[8])染色体,均编码CLV1基因[78~80]。
8 总结
油菜及其近缘植物已定位、克隆的形态标记性状基因列于表1。从表1可见:(1)控制同一性状的基因往往不只1个,而是多个。(2)这些控制同一性状的多个基因可能位于不同代谢途径(如矮秆),也可能位于同一代谢途径(如紫叶)。如果通过基因编辑等手段创制新的种质,可以选择不同途径、不同靶标基因进行研究。(3)异源四倍体的油菜、芥菜控制性状的基因数目一般是其祖先亲本种白菜、甘蓝的2倍,在相同的染色体或同源区段能找到同源基因,这为进一步克隆油菜基因提供了借鉴。
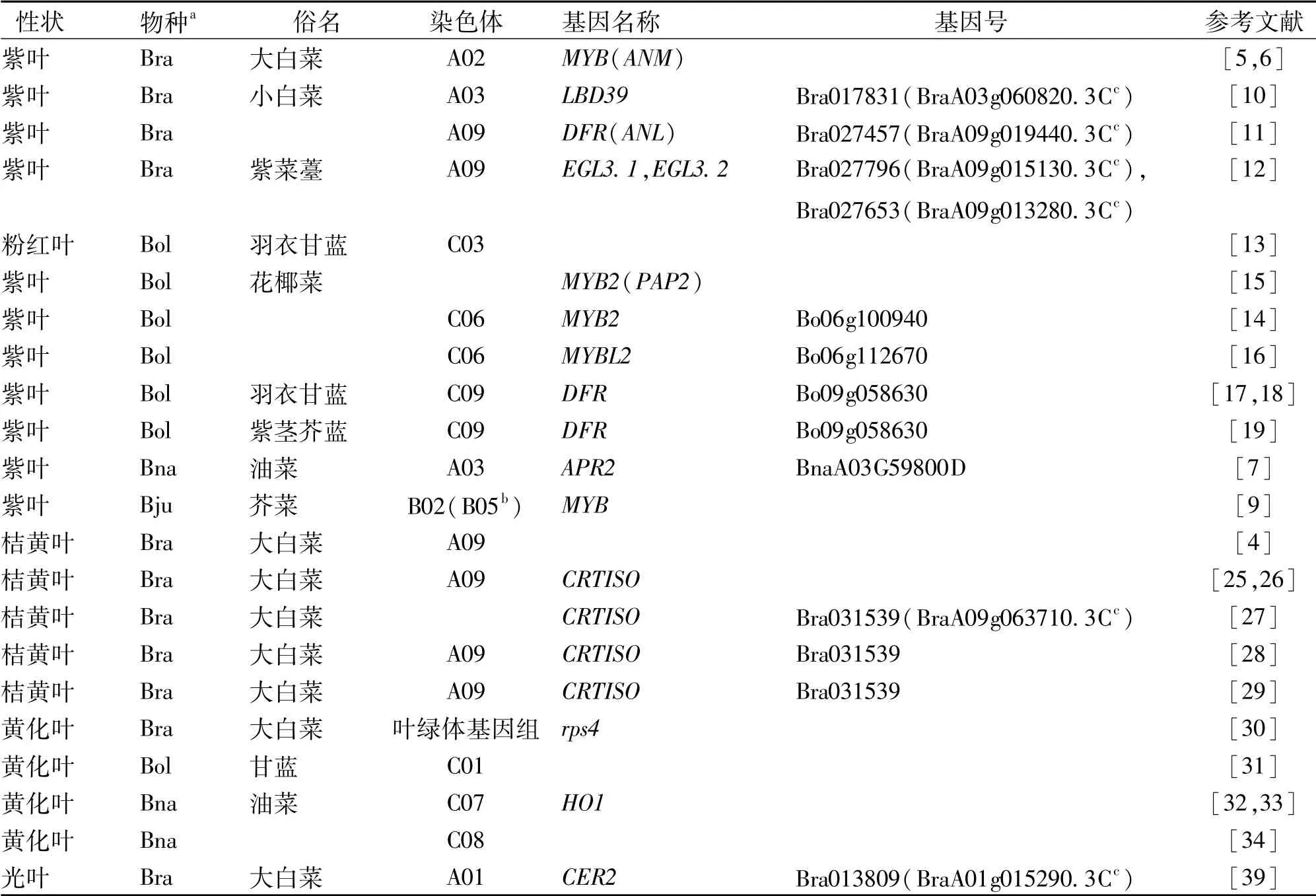
表1 油菜及其近缘植物已定位和克隆的形态标记性状基因

续表1
