Synapsin1磷酸化参与开心散改善Aβ诱发的小鼠记忆障碍*
2019-01-16黄树明徐红丹刘学伟
张 博,黄树明**,徐红丹,刘学伟
(1.黑龙江中医药大学中医药研究院 哈尔滨150040;2.黑龙江中医药大学/佳木斯学院佳木斯154007;3.齐齐哈尔医学院/药学院 齐齐哈尔161006)
阿尔茨海默病(Alzheimer's disease,AD)是一种常见的中枢神经系统退行性疾病,已成为世界生命科学领域的难题之一[1]。β淀粉样蛋白(amyloidβ,Aβ)是AD发病的关键环节[2],其可溶性的寡聚体具有极强的神经毒性并引发一系列级联反应[3-4],但目前尚无切实可行的治疗AD的对策。
开心散(Kaixin-San,KXS)始见于唐代《备急千金要方》,由“远志、人参、茯苓、石菖蒲”组成,具有“主好忘”之功。大量研究证实开心散能够改善动物的学习记忆能力,但目前关于开心散改善记忆障碍的研究多从抗氧化、保护神经元、抑制乙酰胆碱酯酶等角度出发[5],缺乏关键的内在机制的研究,而本实验从内在机制角度出发进行研究,以深入阐明开心散改善记忆能力的作用机制。突触可塑性是学习记忆的电生理学基础,是记忆形成的关键,其形成过程需要突触前膜和突触后膜相关机制的共同参与,即突触前膜递质释放和(或)突触后膜反应的敏感性。我们课题组前期研究证实开心散可改善Aβ诱发的突触可塑性抑制[6],且在突触后膜机制方面,谷氨酸受体2蛋白介导了这一过程[7],但对于突触前膜机制尚未见到相关的报道,而本研究将重点锚定在突触前膜机制上。突触蛋白(Synapsin,SYN)是突触囊泡功能调节的重要蛋白,其是否磷酸化极大的影响突触后膜囊泡递质的释放率,同时递质释放是突触前膜相关受体蛋白发生功能和结构改变的基础,因此为探讨开心散改善突触可塑性抑制的机制,本实验通过观察开心散对模型小鼠行为学,突触前膜递质释放效率及突触前膜磷酸化突触蛋白1(phospho-SYN1,p-SYN1)和 突 触 蛋 白1(Synapsin1,SYN1)蛋白表达的影响,明确开心散影响学习记忆能力的突触前膜分子机制。
1 材料与方法
1.1 实验动物
10 周龄雌性ICR小鼠(30±5 g),购自黑龙江中医药大学,许可证号:SCXK(黑)2008-004。常规饲养在SPF级环境中,温度为24±2℃,湿度为55±5%,12 h昼夜交替。所有动物实验均按照科技部发布的《实验动物福利伦理指南》执行,且经黑龙江中医药大学动物实验伦理委员会批准。
1.2 药物与试剂
人参(Panax ginsengC.A.Mey.)、茯苓(Poria cocos(Schw.)Wolf)、远志(Polygala tenuifoliaWilld.)、石菖蒲(Acorus tatarinowiiSchott)均购自北京同仁堂药店,并由黑龙江中医药大学生药学王喜军教授鉴定。人参、茯苓、远志、石菖蒲按3∶3∶2∶2比例组成。依据课题组前期提取工艺的研究结果,即:以浸出物收率总皂苷为质量控制的主要指标,根据正交设计及方差分析,确定最佳提取工艺为用10倍量的60%乙醇回流提取3次,每次1.5 h,合并提取液,滤过,药液浓缩至相对密度为1.10(60℃)的清膏,减压(真空度为0.06-0.075 MPa)于60℃干燥箱内干燥至恒重,粉碎成细粉,冻存,备用。此工艺条件下浸出物收率为27.6%,总皂苷含量为5.26%。
Aβ1-42(Sigma,A9810),全蛋白提取试剂盒(Solarbio,BC3710),BCA蛋白浓度测定试剂盒(碧云天,P0012S),SDS-PAGE凝胶制备试剂盒(Solarbio,P1200),兔源SYN1一抗(Bioss,bs-3501R),兔源p-SYN1(Ser9)(Bioss,bs-3289R),β-actin(Bioss,bs-0061R),辣根酶标记山羊抗兔二抗(中杉金桥,ZB-2301),超敏ECL发光液(MA0186,大连美仑生物技术有限公司)。
1.3 实验仪器
Morris水迷宫(安徽淮北正华生物仪器设备有限公司),刺激电极(SNEX100同轴双极金属电极,KOPF公司),记录电极,放大器(DAM80,World Precision Instruments),刺激输出装置(SS-104J,NIHON KOHDEN),电刺激隔离器(SEN-80203,NIHON KOHDEN),数据获取装置(Digidata 1440A,Molecular Device),双目万向支架体视显微镜(ZX-0745-1W,深圳市众寻光学仪器有限公司),稳压电源(DJW-1000,上海全力电器有限公司),自动酶标检测仪(Thermo Fisher),Western-blot仪器包括电源,电泳槽,转膜仪及凝胶成像系统均来自BioRad公司。
1.4 模型制备及给药
小鼠随机分为3组,即对照组,模型(Aβ)组和开心散(Aβ/KXS)组。小鼠经戊巴比妥钠45 mg·kg-1腹腔注射麻醉后,固定于小鼠脑立体定位仪上,头部备皮,消毒手术区皮肤,沿头部正中线做1 cm-1.5 cm切口,暴露前囟及人字沟,剥离骨膜。参照《小鼠脑立体定位图谱》[8]定位侧脑室,在前囟后1 mm,中线旁开1.75 mm,用骨钻在颅骨上钻孔,暴露硬脑膜,微量注射器垂直脑表面进针1.8 mm,向单侧脑室缓慢注入5μL Aβ1-42(浓度为500μM的Aβ1-42用生理盐水稀释成浓度为1μM),进针5 min,留针5 min,以防液体溢出。对照组在相同位置注射等体积生理盐水。Aβ及Aβ/KXS组均在侧脑室注射Aβ1-42。Aβ/KXS组小鼠于造模前7天给予开心散灌胃0.2 mL(0.15 g·kg-1),对照组及Aβ组给予等体积生理盐水灌胃,每日一次,持续至电生理学测试当天。因预实验中通过行为学实验对开心散低、中、高剂量组小鼠进行了学习记忆能力检测,发现开心散中、高剂量组小鼠学习记忆能力得到改善,也就是开心散中剂量即为有效剂量,因此本实验选用开心散中剂量,即0.15 g·kg-1的剂量进行实验。
1.5 观察指标
1.5.1 Morris水迷宫实验
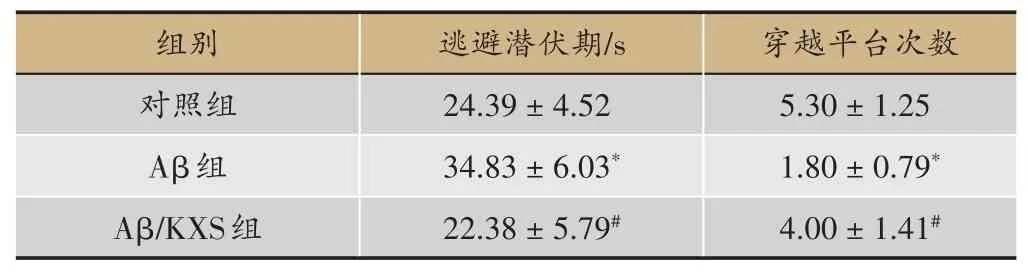
表1 开心散对模型小鼠学习记忆能力的影响(n=10)
造模前、后分别对各组小鼠进行测试,将基础智力和体力不合格者予以剔除。水迷宫测试分为两部分:①定位航行实验:将平台放在第Ⅰ象限中间,在每个象限(4个象限)固定一点面向水池池壁将小鼠放入水中,每个象限各测试一次,每次测试60 s。若小鼠在60 s之内尚未找到平台,则将小鼠引导至平台上并使它在平台上停留10 s,若小鼠在60 s之内找到平台,也让其在平台上停留10 s,方结束一次训练,如此训练5天。第5天训练时小鼠找到水下平台的时间即逃避潜伏期。②空间探索实验:第5天训练结束后,将平台移走,2 h后将每组小鼠从固定象限(第Ⅱ象限)的一点放入水中,记录每只小鼠在60 s内穿越平台的次数。
1.5.2 新奇事物认知实验
小鼠在鼠笼适应5 min后,将2个体积基本相同的不同物体(A、B)放入鼠笼,分别观察并记录小鼠在10 min内对A、B进行探索的时间,计算探索偏好指数a(%)=【(B/A+B)】×100。实验室保持安静,避免强光刺激。24 h后,将物体(A)和新物体(C)放入笼中,分别观察并记录小鼠对A、C进行探索的时间,计算探索偏好指数b(%)=【(C/A+C)】×100。
1.5.3 电生理学测定
将已麻醉的小鼠固定于小鼠脑立体定位仪上,用骨钻分别于刺激及记录电极处钻孔备用,实验中用电热毯维持动物体温于37℃。记录电极采用玻璃微电极,内充满生理盐水,电极尖端内径以电极电阻小于3 M欧姆为准;刺激电极采用同轴双极金属电极。突触后电位的记录选择研究海马突触可塑性常用perforant pathway(PP)—齿状核(dentate gyrus,DG)通路,实验时利用微推进装置将电极插入预定的刺激/记录部位。测定PP-DG通路突触后电位时,电极插入后刺激电极的尖端定位在海马perforant pathway侧枝处,坐标为Bregma后4.5 mm、中线右侧旁开3.0 mm,深度为皮层下1.5-2.0 mm;记录电极尖端定位在DG区分子层,坐标为Bregma后2.1 mm、中线右侧旁开1.5 mm,深度为皮层下1.75-2.25 mm;参考电极固定于后头部皮肤表面。放大器滤过低于10 Hz及高于3 kHz的波。
基础电刺激强度以诱发场兴奋性突触后电位(field excitatory postsynaptic potential,fEPSP)最大振幅的30%作为基准。待基线记录稳定15 min后,给予1串100个脉冲组成的高频刺激(100 Hz,1 sec),然后观察并记录电刺激后第一分钟的fEPSP锋电位(population spike,PS)振幅,数据以PS振幅的百分比表示。
采用Clampex 10.2软件记录fEPSP波形。记录完成后,用Clampfit 10.2软件分析上述记录到的波形获得并处理数据,数据以(±S.E.)表示。
1.5.4 突触前膜p-SYN1和SYN1蛋白含量检测
将前期电生理实验结束冻存的各组刺激侧小鼠海马组织冰上裂解,离心取上清液。BCA蛋白浓度绘制标准蛋白曲线,测定各组小鼠海马组织的蛋白浓度,依据测定的蛋白浓度按比例向各组组织中加入4×上样缓冲液,煮沸,离心。配制8%分离胶和5%浓缩胶,每孔上样20μg制备好的蛋白样品。上层胶70 V,电泳30 min,下层胶110 V,电泳1 h。湿转仪外部置于冰水中,100 V湿转2 h,TBST缓冲液洗5 min×1次。室温封闭液封闭1 h,p-SYN1和SYN1一抗(1∶500)4℃孵育过夜,TBST缓冲液洗5 min×5次。二抗(1:5000)室温孵育1 h,TBST缓冲液洗5 min×5次。ECL发光液室温孵育1min,凝胶成像系统曝光成像并进行体积分析。
1.6 统计学分析
2 结果
2.1 开心散对模型小鼠学习记忆能力的影响
2.1.1 水迷宫实验
水迷宫定位航行试验第5天,记录各组逃避潜伏期和穿越平台次数如图1,实验结果见表1。结果显示,与对照组相比较,Aβ组小鼠逃避潜伏期明显延长(P<0.05),穿越平台次数明显减少(P<0.05),说明Aβ损伤了小鼠的学习记忆能力。与Aβ组相比较,Aβ/KXS组小鼠避潜伏期明显缩短(P<0.05),穿越平台次数明显增加(P<0.05),说明开心散改善了小鼠的学习记忆能力,提示开心散可改善Aβ诱发的记忆障碍。
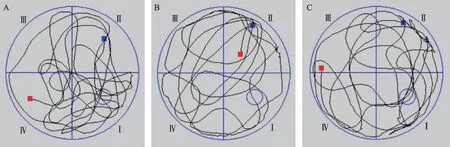
图1 各组小鼠水迷宫空间探索实验运动轨迹示意图
2.1.2 新奇事物认知实验
新奇事物认知训练当天及24 h后,记录各组小鼠探索偏好指数,分别为探索偏好指数a及探索偏好指数b,实验结果见表2。结果显示,训练当天Aβ组与对照组的探索偏好指数a及Aβ/KXS组与Aβ组的探索偏好指数比较,均无统计学差异(P>0.05)。24小时后,放入新物体C后,与对照组比较,Aβ组小鼠对C物体的探索偏好指数b明显降低(P<0.05),即Aβ组小鼠对物体A和C的探索时间接近。而与Aβ组比较,Aβ/KXS组小鼠探索偏好指数b明显增加(P<0.05),即Aβ/KXS组小鼠对物体C的探索时间明显增加。说明Aβ损伤了小鼠的学习记忆能力,而开心散改善了Aβ所致的记忆损害。
2.2 开心散对模型小鼠突触前膜递质释放效率的影响
fEPSP波形稳定基础记录15 min后,给予一串高频刺激所诱发PS振幅增加的第一分钟,间接反映的就是突触前膜递质释放效率,见表3。电生理学实验结果显示,与对照组(n=20)相比,Aβ组小鼠PS振幅降低(P<0.05),与Aβ组(n=6)相比,Aβ/KXS组(n=7)小鼠PS振幅明显增加(P<0.05),说明Aβ抑制了突触前膜递质释放效率,而开心散可改善Aβ对突触前膜递质释放效率的抑制作用。
2.3 开心散对模型小鼠海马突触前膜蛋白SYN1及p-SYN1表达的影响
Western-blot结果显示,Aβ组小鼠海马p-SYN1表达下降,且p-SYN1/SYN1灰度比值为0.31±0.04,与对照组(0.56±0.09)比较,数值明显降低(P<0.05),说明Aβ抑制了SYN1磷酸化,提示磷酸化的SYN1(Ser9)可能参与了Aβ对小鼠海马突触可塑性的抑制。Aβ/KXS组小鼠海马p-SYN1表达相对增多,且p-SYN1/SYN1灰度比值为0.58±0.05,与Aβ组相比较,比值明显升高(P<0.05),说明开心散增加了SYN1的磷酸化,且逆转了Aβ所致的p-SYN1/SYN1的比值下降,提示开心散可能通过促进SYN1磷酸化进而改善Aβ诱发的突触可塑性抑制从而改善学习记忆能力(见图2)。而在图中SYN1显示的双条带可能与其具有至少2个亚型有关。

表2 开心散对模型小鼠学习记忆能力的影响(n=10)
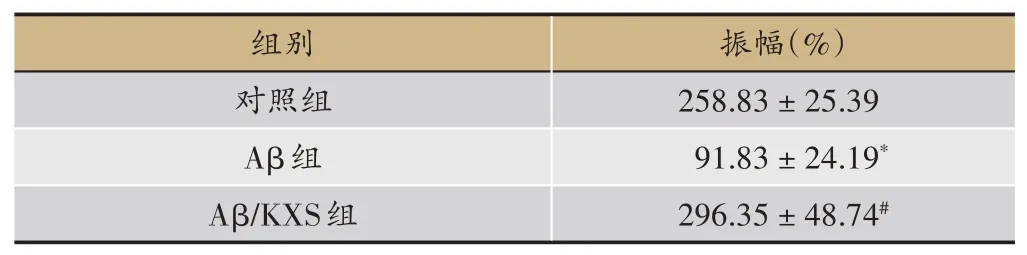
表3 开心散对模型小鼠突触前膜递质释放效率的影响
3 讨论
AD患者的临床表现之一即记忆力减退,而Aβ是其发病的关键,具有神经毒性,严重损害患者记忆和认知功能。目前建立AD动物模型的方法有很多包括转基因动物、Aβ脑内注射等[9-10,11,12],以模拟与AD患者病理表现相似的脑内Aβ聚集,进而研究AD的发病机制。截至目前,通过复制AD模型动物已有大量行为学实验结果证实Aβ可损伤AD模型动物的学习记忆功能[13],并通过多个环节起作用[14-15],而Aβ脑内注射是目前研究最多,应用最广泛的AD动物模型造模方法,故本实验采用Aβ脑内注射的造模方法复制AD动物模型。本实验通过两种行为学检测方法共同证实,在基础智力无差异的情况下,Aβ所致AD模型小鼠的学习记忆能力较对照组小鼠明显下降,表明Aβ能够损伤学习记忆能能力,该实验结果与文献报道相一致。

图2 突触前膜p-SYN1及SYN1蛋白表达的Western-blot结果
中药复方具有多因素,多靶点,多途径的作用特点,因此在治疗AD上具有独特的优势。经方开心散中以人参、茯苓,大补元气,滋补脾肾,淡渗利湿,维护先后天之本;远志、菖蒲安神定志,化痰祛邪以开窍,从而达到益气扶正,化痰祛邪的治疗目的。临床研究发现开心散变化方能有效改善AD患者认知功能方面的临床症状[16],且大量实验通过行为学研究发现开心散具有改善模型动物记忆障碍的作用[9-10,17],本研究通过多种行为学方法亦证实在各组小鼠基础智力无差异的情况下,Aβ/KXS组小鼠的学习记忆能力较Aβ组小鼠有所改善,说明开心散可改善Aβ诱发的小鼠记忆障碍,这与前期研究结果相一致[7]。
尽管大量实验表明开心散可提高AD模型动物记忆力,且可通过多途径治疗AD,但关于开心散对突触信息传递功能可塑性的影响报道较少,仅日本学者于1994年报道开心散可促进大鼠海马齿状核长时程增强(long-term potentiation,LTP)的形成,从而改善乙醇所致记忆障碍模型大鼠学习缺陷[18],而其也证明开心散复方各组分药物中人参和茯苓起到了增强海马LTP的作用[19]。但开心散对AD模型动物LTP的影响及其影响LTP的机制未见报道,然而我们课题组前期研究证实开心散可改善AD模型动物突触可塑性[7]。
突触之间信息传递主要是由突触前末梢突触囊泡所释放的递质介导,SYN是突触囊泡中含量最为丰富的跨膜蛋白,其含有多个亚型,其中SYN1在突触小泡移动方面起主要作用,同时SYN也是突触囊泡的特异性标志蛋白,是突触囊泡功能调节的最重要的分子之一并参与递质释放[20]。通常情况下突触囊泡由突触蛋白锚定于细胞骨架丝上,当轴浆内Ca2+浓度升高时,引发一系列反应,促使突触蛋白Ser9位点发生磷酸化,进而与骨架丝结合力减弱,突触囊泡游离,促进递质释放[21-22]。递质大量释放后,相关递质与突触后膜受体结合,启动突触后膜机制,增强突触可塑性[23]。突触可塑性是学习记忆的电生理学基础[24],突触可塑性的发生主要涉及突触前膜机制和突触后膜机制,课题组前期的研究结果发现在开心散改善Aβ诱发的突触可塑性抑制的过程中,突触后膜相关机制起到重要作用[7],但突触前膜机制并不清楚。因此本实验通过电生理学方法间接测定了突触前膜递质释放效率,如果开心散对突触前膜递质释放效率有影响,那么本研究将采用Western-blot法重点检测突触前膜p-SYN1(Ser9)和SYN1蛋白表达的改变。
实验结果显示在给予高频刺激第1分钟,即强直后增强期,Aβ组小鼠PS振幅明显降低,说明Aβ抑制了突触前膜递质释放,而开心散组小鼠PS振幅明显升高,说明开心散能够改善Aβ对突触前膜递质释放效率的抑制。因此我们进行了Western-blot实验,结果显示Aβ组小鼠海马p-SYN1表达降低,且p-SYN1/SYN1比值明显下降,说明Aβ降低了突触前膜SYN1的磷酸化水平,这与前人研究结果不一致,可能与在体实验的影响因素较复杂有关[25]。而给予开心散灌胃的小鼠海马p-SYN1表达增加,且p-SYN1/SYN1比值升高,说明开心散能够增加SYN1的磷酸化水平,逆转Aβ诱发的p-SYN1/SYN1比值下降,提示开心散可能通过促进SYN1磷酸化,增加递质释放,从而减轻Aβ所致的突触可塑性抑制。同时,实验发现Aβ/KXS组小鼠海马SYN1表达水平较Aβ组减少,这可能与开心散可以降低Aβ引发突触前膜递质释放紊乱带来的损害有关[25]。
综上,开心散通过增加SYN1磷酸化蛋白表达进而改善Aβ诱发的突触可塑性抑制,可能是开心散改善Aβ所致小鼠记忆障碍的机制之一。然而,突触前膜含有多种涉及到突触传递的因子,机制也较为复杂,本研究仅发现突触前膜的SYN1及其磷酸化参与开心散改善Aβ所致突触可塑性抑制,但详细的突触前膜机制有待进一步研究。
