越冬条件对豚草种子内生真菌寿命和种子发芽能力的影响
2019-01-14王国红李绍锋杨民和张斯琴张碧尧
王国红,李绍锋,杨民和,张斯琴,张碧尧
(福建师范大学生命科学学院/细胞逆境响应与代谢调控福建省高等学校重点实验室,福建福州350108)
豚草(AmbrosiaartemisiifoliaL.)是起源于北美的一年生菊科植物,种子是唯一的繁殖体,也是扩散的唯一途径。近200年来,豚草迅速在世界各地蔓延,在美洲、欧洲、亚洲和澳洲等地区均有分布[1-2]。豚草具有广泛的生态适应性和很强的竞争能力,是一种严重危害农业、生态和人体健康的恶性杂草。豚草自20世纪30年代侵入中国,发生范围迅速扩张,目前在我国21个省、市和自治区广泛分布[3]。在福建省,豚草于1993年首次在长乐市被报道[4]。
针对生物入侵的机制,学术界陆续提出了一些假说,其中比较重要的有天敌逃避假说[5]、入侵种的引入后进化假说[6]、促进的共生假说[7]等。豚草种子特性,如种子数量大、发芽的生态可塑性、种子初次休眠和再次休眠以及土壤种子库是决定其生态适应性的关键因素[8-9]。同时,豚草的生态可塑性很强,其种子发芽和休眠特性随分布纬度的不同而变化,分化出适应当地环境条件的生理种群[1-2]。进入新的地域后,入侵植物与生物的相互作用是影响种群建立和扩散的另一个关键因素[7,10]。真菌是土壤中和植物体的常见寄居者,可作为病原菌导致种子、芽和幼苗的腐烂[11],或作为益生菌促进种子生活力和生长[12],从而影响植物的生态适应性和竞争力[13]。豚草种子成熟后落入土壤,不可避免地要和土壤生物(包括真菌)发生关系[14]。内生真菌是一类生活于健康植物体内,对植物没有引起病害症状的微生物。豚草种子中生长有丰富的内生真菌,一些真菌的发酵液对豚草种子发芽和幼苗生长有不同的生物学作用[15]。
中国的福建省长乐市冬季气候温暖湿润,与豚草的北美原产地差异明显[4]。豚草扩张进入福建省沿海地带后,温暖湿润的冬季环境,以及不同的越冬条件可能对豚草种子休眠、发芽和种带内生真菌等产生影响。本研究通过分析豚草种子越冬和储存期间种子内生真菌的存活能力和种子发芽能力的动态变化;土壤种子和植冠种子内生真菌感染和种子发芽的动态变化;以及实验条件下土壤种子的发芽动态,旨在考察温暖冬季和不同越冬条件对豚草种子内生真菌存活与种子发芽能力的影响,探讨越冬条件、种子内生真菌和豚草种子发芽之间的相关性。
1 材料与方法
1.1 研究地域概况
研究区域位于中国福建省长乐市松下镇寨下村(25°46′32″N,119°37′14″E),属于中国东部沿海,是中亚热带和南亚热带的过渡区;气候类型为亚热带海洋性季风气候,温暖湿润,四季分明;年平均温度在15.6~19.3 ℃,积温为6 375.6 ℃,一般7月气温最高(平均温度25~34 ℃),1月最低(平均温度8~15 ℃);雨量充沛,年平均降水153 d,平均降雨量为1 200~1 700 mm,降雨多在3—9月,4—6月为梅雨季节;无霜天数333 d;光照充足,年均日照时数为1 837.6 h,且光照、温度和降水配合良好,雨热同季。土壤为海滩沙土。8—10月台风发生较为频繁,秋、冬季节以东北风为主;风卷沙土,有利于豚草种子的短距离扩散。
1.2 种子采集
选择人为活动较少、接近自然状态的区域为采样点,参照李绍锋等[15]的方法取样。采集植冠种子时,每一采样点随机选取5植株,每一植株采集3个分枝上的种子,将每一样点共25植株种子混合,用透气袋装好,存放在阴凉处备用;每年12月以后,植冠残存的种子逐渐稀少,则将枯枝上所有种子收集。土壤种子的采集,设置5个采样点,每一采样点随机选取1棵豚草植株,以植株为中心、半径为45 cm范围内采取植株底下表面0~5 cm的沙土,用16目的筛子筛去沙土,挑出豚草种子,用透气袋装好,编号后存放于实验室阴凉处备用。为系统分析豚草种子发芽能力的变化,在种子成熟后,于每月底采集土壤和植冠上的豚草种子,分离种带真菌,检测种子的发芽能力。
1.3 不同的种子越冬处理
分别采用3种方法储存豚草种子,经历一定储存时间的种子用于内生真菌的分离或是发芽试验。(1)室内透气袋储存:将当年采集的新鲜植冠豚草种子装入透气袋中,置于室温条件下储藏。(2)野外沙土埋藏:2013—2015年,每年10月采集豚草种子用于野外种子埋藏试验。在收集种子的同时,采集豚草生长地沙土,该沙土经过16目的筛子过筛,检查无豚草种子残留后用于试验;50粒种子混入500 g沙土中,用尼龙网袋装好,放入豚草生长地表层土壤中(0~5 cm),使其经历越冬;选择有枯死豚草植株的5个埋藏点,每一埋藏点间隔至少0.5 m,至次年3月收集经历越冬的种子,用于试验。(3)室内沙土储存:挑选健康、颗粒饱满的种子400粒,放置于装有豚草发生地沙土的花盆表层(0~5 cm),花盆放置于实验室内。以上3种方式越冬后的种子与在豚草发生地植冠上的种子、沙土中自然越冬的种子一起,用于分离内生真菌,比较不同越冬方式对种子内生真菌分离率和发芽的影响。
1.4 种子内生真菌的分离
豚草种子先用清水清洗以去除杂质,浸泡过夜后沥干水分。豚草种子的表面消毒程序参照李绍锋等[15]的方法:先用体积分数75%乙醇漂洗30 s,再经3%次氯酸钠浸泡8 min,后在体积分数75%乙醇溶液中漂洗30 s;消毒剂处理后的种子用无菌水洗涤3次,每次5 min,以漂洗除去残留的消毒剂。然后在超净工作台中适当晾干水分后接种于PDA平板上,每皿7~9粒。接种完成后,将培养皿用透气带封好,置于(28±1)℃恒温下培养。待有真菌长出时,逐个挑出移接到新的平皿上进行纯化、鉴定,记录内生真菌的分离率。种子内生真菌分离率表示为培养后生长出真菌的种子数占分离用种子数的百分率。
1.5 野外调查、试验
每年11月下旬开始,即可见枯死的豚草植株下有大量的豚草幼苗发生。采用5点取样法,每个样点选择1株枯死豚草,以植株为中心,45 cm为半径,采集植株下方深度为0~5 cm范围内的沙土及所有幼苗,用16目的筛子筛去沙土,计算幼苗数和种子数,未发芽的种子继续用于发芽试验。
2014年3月下旬,在福建省长乐市松下镇豚草自然发生地发现3株处于开花期的豚草植株,此后定点、定期观察植株的生长情况。于2014年5月7日收获种子165粒,其中饱满成熟种子97粒,瘦小种子68粒;取50粒饱满种子用于发芽试验。
1.6 种子萌发试验
滤纸保湿法:将培养皿底部垫上滤纸,每皿50粒种子,清水保持滤纸湿润,于温度为28 ℃光照培养箱中萌发;箱内湿度为70%,光周期(L ∶ D)为12 ∶ 12。记录发芽种子数,计算发芽率,种子出现胚根即认为已经萌发;同时记录种子腐烂、休眠和死亡等情况。种子发芽率表示为发芽种子数占试验种子数的百分率。每个处理设置3个重复。
花盆沙土培养法:豚草发生地沙土经16目筛子过筛,除去其中的植物种子和其他杂质,均匀地将沙子分装到5个同等大小的花盆中,用适量的清水浇湿沙土;分别将种子放置到对应花盆沙土表面,尽量模拟自然条件,每隔2 周计算发芽种子数,并将长出的幼苗拔除。发芽处理结束后,用16目的筛子筛出残余种子,观察分析种子的腐烂、休眠和发芽等情况。
1.7 种子生活力测定
种子生活力采用TTC (2,3,5-triphenyl tetrazolium chloride,TTC) 染色法检测[11]。种子充分吸水膨胀后,用解剖刀剥离果皮,放入1.0%的TTC染色液中,以浸没为度,置于28 ℃的恒温培养箱中,黑暗中进行染色反应8 h;染色完成后倒去染色液,用清水冲洗种子数次,然后观察染色情况,判断种子是否有生活力。
1.8 数据统计分析
采用SPSS19.0统计软件对试验数据进行统计分析,Origin 9.1软件作图,不同处理间参数的差异在0.05水平上采用Duncan法进行比较。
2 结果与分析
2.1 室内储存种子内生真菌发生的动态变化
室内透气袋储存条件下,豚草种子经不同时间的储存后,随着时间的增长,种子内生真菌的分离率呈现逐步下降的趋势,结果见图1。10月刚成熟的豚草种子真菌分离率为78.14%,分离获得的真菌种类较为丰富(结果未显示);存放3个月后,内生真菌分离率下降至42%;随着储存时间的持续延长,内生真菌的分离率明显降低。存放9个月后,内生真菌的分离率仅为8%,与储存3、6个月种子的真菌分离率相比,达到差异极显著水平(P<0.01)。储存12个月的种子难于分离到内生真菌。2010年10月从长乐采集的种子也得出类似的结果,种子存放15个月后即难于检测到内生真菌(结果未显示)。
2.2 不同方式越冬后种子内生真菌的变化
不同条件越冬后豚草种子中内生真菌发生情况的比较见图2。从图2可知,自2013年10月至2014年3月底(约为种子成熟后6个月),自然环境中越冬的豚草种子中,植冠上种子的内生真菌分离率为73.16%,明显高于沙土中种子(61.93%)(P<0.01)。人为用沙土覆盖的种子越冬后,其内生真菌分离率分别为56.90%和65.88%。以上处理的种子其内生真菌分离率均明显高于室内透气袋保存的种子(20.07%)(P<0.01)。总体来看,内生真菌的分离率由高到低依次为植冠种子(73.16%)、沙土中种子(56.90%~65.88%)和室内透气袋保存的种子(20.07%)。

柱状图上不同字母表示数值在0.05水平上差异显著(下同)Different letters on the column show significant difference at the 0.05 level (the same below)图1 豚草种子储存期内生真菌的动态变化(均值±标准误)Fig.1 Variation of isolaton rates of endophytic fungi from common ragweed seeds during the storage (average±SE)

图2 不同越冬方式对豚草种子内生真菌分离的影响(均值±标准误)Fig.2 Difference of isolation rate of endophytic fungi in common ragweed seeds after over-winter periods (average±SE)
2.3 自然条件下越冬期种子内生真菌变化动态
2014年,自豚草种子成熟的10月底起,逐月定期到长乐采集豚草植冠上和沙土中的种子,调查越冬期种子内生真菌的变化动态,结果见如图3。10月份刚成熟的种子均存在于植冠上,沙土中种子稀少。11月后,落入沙土中的种子逐渐增多。11月份,植冠上和沙土中的种子其内生真菌分离率相当(P>0.05);随着越冬时间的增长,沙土中的种子内生真菌分离率明显降低(P<0.01),其中12月底内生真菌的分离率最低为38%。内生真菌的种类也发生较大的变化,新鲜的种子主要以链格孢属真菌为主,随着与土壤接触时间的增长,来自土壤中的真菌如青霉菌和木霉菌比例增多。植冠上种子在越冬期内真菌分离率较为稳定,其中1月底内生真菌分离率最低为63.78%,其它月份内生真菌的分离率均在70%以上,真菌种类上主要以链格孢属和镰刀属的真菌为主。总体上,落入沙土中2个月后,沙土中的种子其内生真菌分离率显著低于植冠种子(P<0.01)。
2.4 不同越冬方式对种子发芽的影响
野外调查发现,在长乐市松下镇寨下村豚草产地,最早于4月下旬即可有种子成熟,于2014年5月7日采获的165粒新鲜种子(部分种子的总苞尚为青绿色),经室内发芽试验,发芽率可达41.18%。
自种子成熟至次年3月,经历越冬以后残留于植冠上的种子其发芽能力最强,可达58.11%(图4);野外人工埋藏的种子发芽率为53.94%,达到差异显著水平(P<0.05)。室内沙土埋藏、室内透气袋储存和野外自然残存于沙土的种子,其发芽能力则大幅度下降,比如经历6个月的室内透气袋存放后,发芽率仅为22.14%。
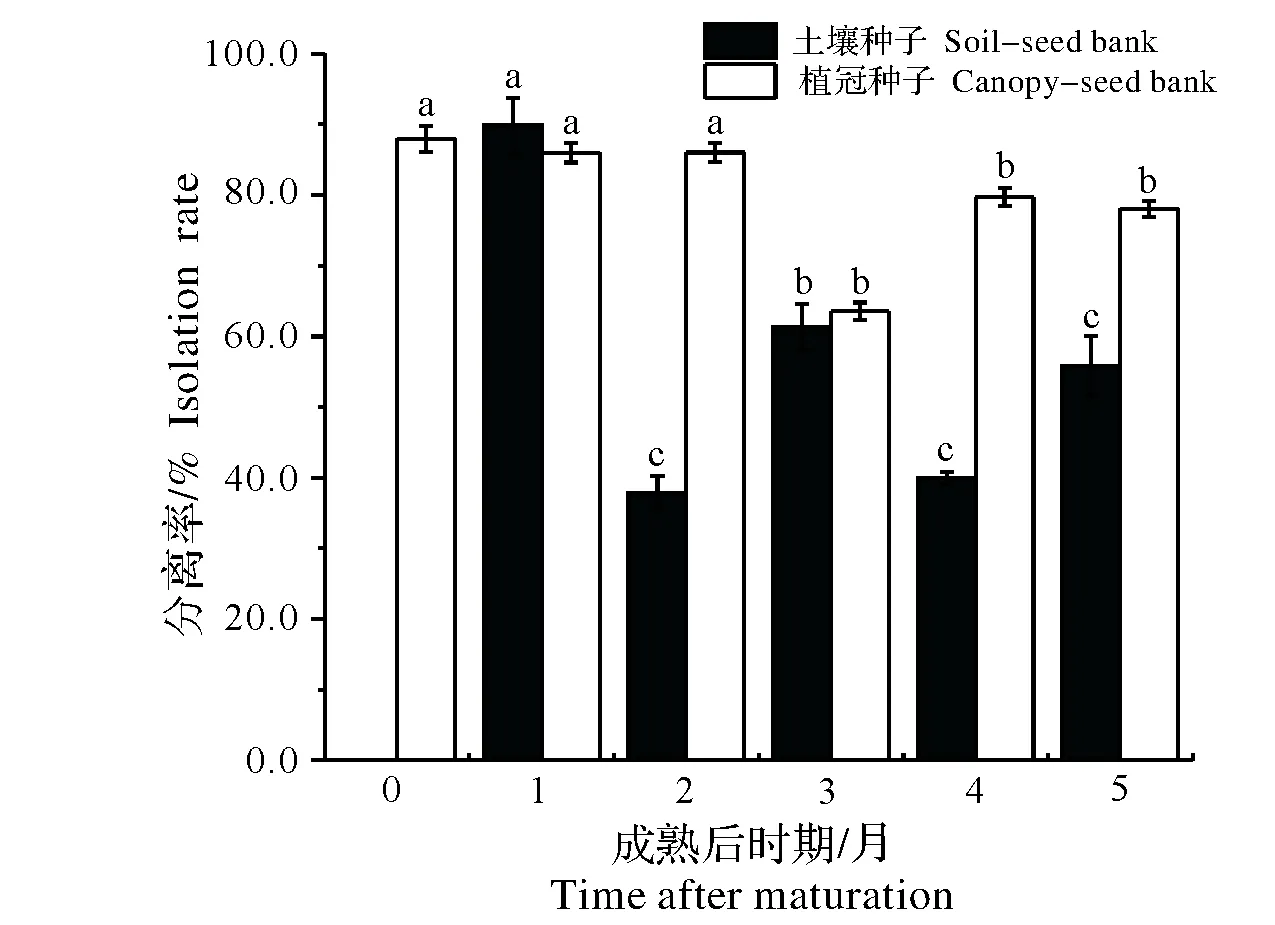
图3 越冬期野外豚草种子内生真菌的变化(均值±标准误)Fig.3 Dynamics of isolation rate of endophytic fungi in common ragweed seeds during the over-winter stage (average±SE)
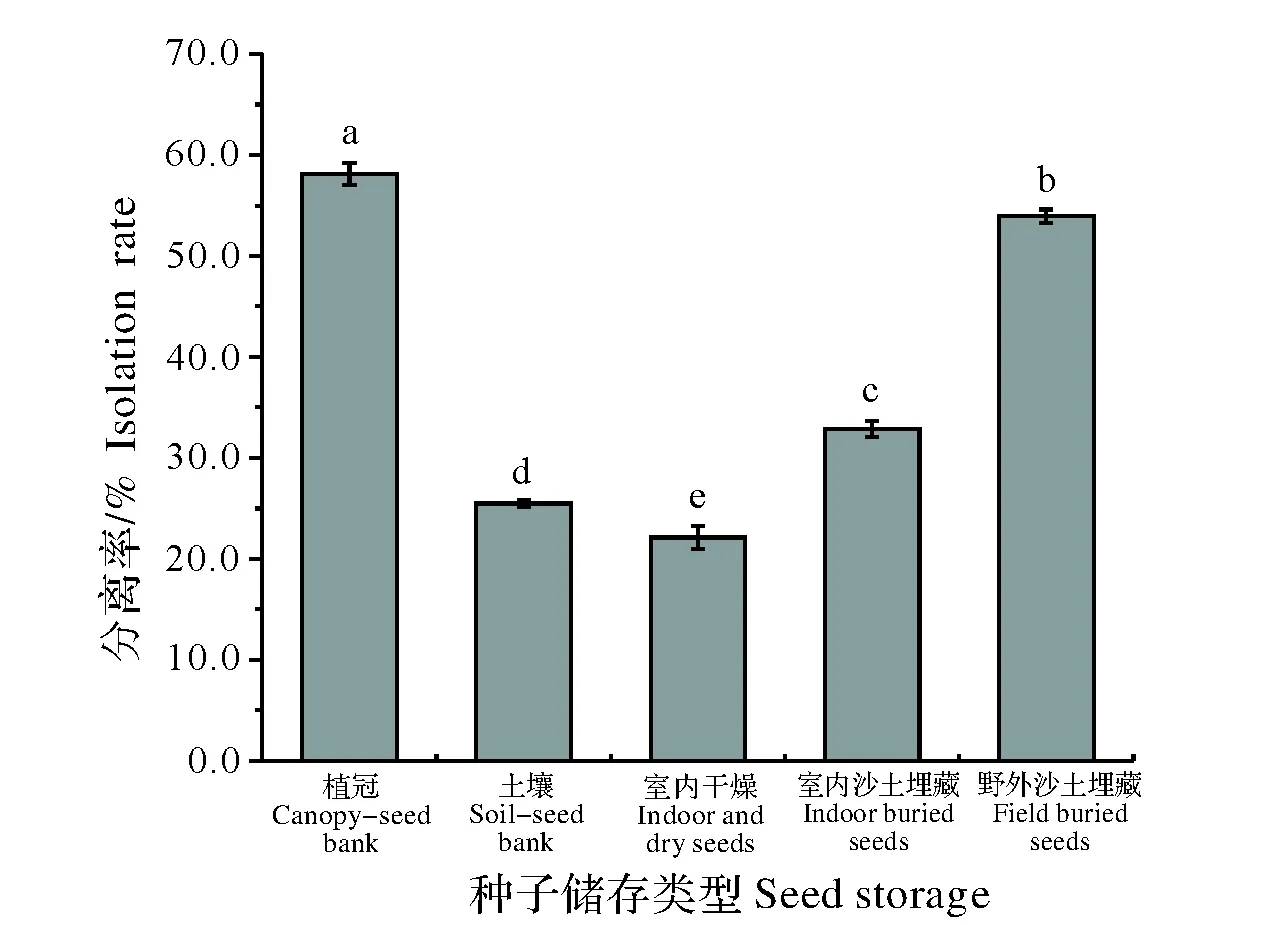
图4 不同越冬方式对豚草种子发芽的影响 (均值±标准误) Fig.4 Effect of over-winter patterns on seed germination of common ragweed (average±SE)
2.5 实验条件下土壤种子萌发动态
2015年11月从5个取样点获得种子2 776粒,将这些种子洒落在沙土表面,置于室外培养,观察种子出苗情况。在近4个月的发芽试验过程中,种子发芽随时间的变化情况如图5所示:在越冬的前期种子萌发较多,两周内的发芽率达到20.87%,明显高于其他时间段(P<0.05);随着越冬时间的延长,种子的发芽率总体上呈下降趋势。3个月后,绝大多数的种子已经发芽;至2016年3月28日,经过近4个月的萌发,种子总发芽率达到70.55%。未发芽的种子经TTC染色法检测,27.89%的种子发生腐烂,仅有1.56%的种子保持生活力。

图5 越冬期豚草种子萌发动态(均值±标准误)Fig.5 Dynamics of seed germination of Ambrosia artemisiifolia during the over-winter stage (average±SE)

图6 越冬期豚草土壤种子和植冠种子发芽能力比较(均值±标准误)Fig.6 Comparison of germination rate of soil seed and canopy seed of Ambrosia artemisiifolia during the over-winter stage (average±SE)
2.6 野外条件下豚草种子发芽的动态变化
野外定期采集落入沙土中种子和保留在植冠上的种子,调查越冬期内不同时期种子萌发能力的动态变化(图6)。调查结果表明:当年成熟的豚草种子保持较高水平的发芽能力;当年10月份至次年1月份(种子成熟后4个月),无论是落入土壤中的种子还是残留在植株上的种子,发芽率均保持在40%以上。10月份采集的植冠种子发芽率最高,为86.49%,与12月(种子成熟后3个月)采集的植冠种子差异不显著(P>0.05),与其它月份植冠种子之间发芽率差异显著(P<0.05);至次年3月(种子成熟后6个月),虽然植冠种子的发芽率明显下降(P<0.05),但仍然维持较高的发芽率(58.01%)。落入沙土中的豚草种子,越冬前期(当年11月至次年1月)维持较高的发芽率(55.90%~78.05%);但是,随着越冬期的延续,残存土壤种子的发芽率显著下降(P<0.05);至次年3月,沙土中种子发芽率仅为22.50%。经历1—2月份的低温时期后,植冠种子的发芽率明显高于土壤种子(P<0.01)。
2.7 豚草幼苗内生真菌检测
室内种子萌发获得的幼苗其真菌分离率为0,豚草自然发生地豚草幼苗真菌分离率为65.52%,差异极显著(P<0.01);而且,从菌落形态和菌体形态的观察可知,自然发生地豚草幼苗中分离获到的真菌与种子内生真菌的种类大多数不同。
3 结论与讨论
豚草种子在不同条件下越冬,其内生真菌的存活能力存在明显的差异。在室内干燥条件下,豚草种带内生真菌的分离率随储存时间延长而下降,说明室内干燥的环境不利于内生真菌的存活。种子成熟后,内生真菌存活能力及其对种子生活力的影响的研究在草坪草和牧草种子中研究较多,但研究结果随植物种类、真菌种类、菌株和生境条件不同而有差异[16-17]。室内储存的种子随着存放时间的延长,内生真菌逐渐死亡,真菌的死亡速度快于宿主种子的衰败速度;但在土壤(如土壤埋藏试验)条件下,宿主种子和内生真菌的寿命是相似的[18]。种子从植株上落到土壤表面后,种子的存活率大为上升,但种子中内生真菌的存活则大为下降[19]。Sowley 等[20]研究发现,莴苣(Primulapolyantha)种子经过4 ℃的冷藏后,种子携带的灰霉菌(Botrytiscinerea)分离率下降,并影响种子发芽后植株中灰霉菌的感染程度[20]。以上的研究结果说明种带内生真菌随种子室内储存时间延长而死亡增多和干燥不利于种带真菌存活,本文的研究结果支持这一结论。但在野外条件下,无论是土壤种子还是植冠种子,其内生真菌的分离率均维持在比较高的水平,与室内干燥储存的种子相比差异显著(P<0.05),说明自然条件下的沙土埋藏有利于真菌存活。与土壤种子相比,植冠种子中内生真菌分离率保持较高的水平(>60%),而土壤种子中内生真菌的分离率总体上呈下降趋势,不同月份之间有所波动。野外条件下种带内生真菌分离率不同月份之间出现的波动,可能与土壤真菌的活动、沙土干燥程度和种子取样方法有关,需要作更深入的研究。自然条件下,掉落到土壤中的种子将受到土壤微生物的影响[11-12,14]。但在北美,研究结果显示土壤真菌对落入土壤的豚草种子影响并不明显[12,21]。和北美相比,福建省长乐市区域属于亚热带季风气候,有利于微生物如土壤真菌的繁殖和生长。因此,豚草种子、内生真菌和土壤微生物如何相互影响,需要作进一步的分析。
豚草属于典型的短日照植物。在北美原产地,当年成熟的豚草种子不能发芽,必须经历低于5 ℃,时间在8周左右的低温层积(休眠),种子才能在晚春或初夏(4月下旬—5月上旬)发芽出苗[1-2,22]。然而,豚草种子发芽和休眠特性随分布纬度的不同而变化。如在我国辽宁省沈阳市和黑龙江省牡丹江市,当年成熟的种子不能发芽,全部进入休眠[23-24];而在江西省南昌市,当年成熟的种子有10%没有进入休眠,部分豚草可以冬季前发芽,以幼苗越冬[25];在广东省佛冈县,豚草种子的休眠性弱,当年成熟的种子在25℃下12 d的发芽率达到64.4%[26]。Sang等[27]采集自中国黑龙江省牡丹江市、江苏省南京市和江西省南昌市等3个地区的豚草种子,在5~40 ℃的范围内均能发芽,种子发芽对渗透压、pH值和光照等环境因素的适应范围广。但是,在5 ℃条件下,采自牡丹江市的种子其发芽率明显高于其它两个地区的种子,发芽的最适温度也不一样,这表明3个地区的种子发芽对温度要求产生了分化[27]。豚草种子发芽的最低温度为4.88 ℃,最高温度为42.92 ℃,而诱导豚草种子进入二次休眠的最低温度约为20 ℃;日照长度低于14.5 h时开始进入生殖生长,霜冻(低于0 ℃)导致植株死亡[1,22,27]。发生于福建省长乐市的豚草种子大多数不需经历休眠即可发芽,而且成熟的豚草种子在当年绝大多数(>70%)已经发芽,以幼苗形态越冬,残留于土壤中的活种子很少(约1.56%)。相比于沙土中的种子和植冠上的种子,干燥储存的种子发芽率显著下降(P<0.05)。豚草种子发芽时间不齐,发芽持续的时间很长(约4~6个月)。福建省东南沿海地区(如长乐市松下镇),冬季的温度、光周期和无霜期天数等气候条件均满足豚草生长发育的最低要求,这些气候条件的满足,可能是导致豚草周年发生的原因。发生于福建省长乐市的豚草,与原产地北美(如加拿大和美国)[1,22]和入侵地欧洲(如德国和匈牙利)[28-29]相比,其种子休眠、越冬和发芽等生物学特性已经发生明显的变化。
发生于福建省长乐市的豚草维持一定数量的植冠种子;与室内干燥储存种子和土壤中的种子相比,经历越冬后豚草的植冠种子维持较高的发芽率。植冠种子库是指所有宿存于植冠上的成熟种子形成的种子库。由于每次只掉落几粒种子,使得种子传播时间延长,能持续补充土壤种子库,提高种子的繁衍和扩散效率。同时,植冠种子由于不接触土壤,因而大大减少土壤侵蚀、腐烂性微生物和小型动物的破坏,也延长种子寿命,提高种子成活率,使得植物能够适应不确定环境,因此在荒漠和火烧生态系统中受到广泛的关注[30]。豚草土壤种子库的研究资料非常丰富[1,31-32],但有关豚草植冠种子库的研究未见报道。由于大多数生活力强的种子在当年即已发芽,在经历较为寒冷的冬季(1—2月)以后,残存于土壤中的种子发芽能力大为下降,而植冠种子的发芽能力仍然强劲,与土壤种子相比差异显著(P<0.05)。豚草的植冠种子库对维持自然种群和促进入侵可能发挥重要的作用。
豚草种子实质上是一种瘦果,由坚硬、致密的果壳(总苞和苞片)包裹胚(种子),对胚起到保护作用,促进胚的休眠[31]。豚草种子(瘦果)的内生真菌主要存在于总苞和苞片中,胚中相对较少[15]。本研究中,由种子发芽获得的幼苗内生真菌的分离率为0,说明种子中的真菌并没有垂直传播至幼苗。发生于福建省长乐市的豚草种子休眠能力大为减弱,当年发芽率提高,除了地理和气候的影响之外,是否也和内生真菌有一定的关系[12,33-34],需要作进一步的分析。
