高寒草甸不同演替阶段优势种叶片光合性状研究
2019-01-09李俐蓉刘旻霞焦骄车应弟
李俐蓉,刘旻霞,焦骄,车应弟
西北师范大学地理与环境科学学院,甘肃 兰州 730070
光合作用是植物生长的物质来源和能量来源,也是全球碳循环和其他物质循环的重要环节(李征珍等,2017;管铭等,2014)。与根、茎、种子相比,叶片对水分、温度、光照等环境因子的变化最敏感,是植物进行光合作用的主要器官(Martin et al.,2009;Violle et al.,2007)。常见的叶片光合性状包括:稳定碳同位素(δ13C)、净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、气孔导度(Gs)、叶片含水量(LWC)及叶绿素(SPAD)等,这些生理指标的变化不仅能体现植物的生长策略和资源利用方式,还能揭示植物对不同生长环境的适应(Kazakou et al.,2006;李善家等,2013)。例如,与生长在干旱生境中的植物相比,生长于水分充足的生境中的植物具有更高的LWC、Pn和Tr,这与植物自我平衡调节机制有关,为了适应环境的变化,植物有伸缩性地控制水分散失和提高生长速率(任青吉等,2015)。植物光合性状除在不同生长环境中表现出差异外,不同生长类型之间也存在较大差异(Lawlor,1995;刘旻霞等,2017)。李宏林等(2012)关于青藏高原高寒沼泽湿地在退化梯度上的植物群落变化对湿地水分状况的影响研究发现,相比于禾草类植物,杂草类具有较低的WUE,较高的 Pn,表明这些差异与植物的生长策略和资源利用方式有关。
青藏高原有“地球第三极”之称,平均海拔在4000 m以上。其中,高寒草甸是青藏高原生态系统中最具代表性的植被地理单元,约占青藏高原可利用草场的50%(白炜等,2011)。从20世纪90年代起,寒草甸退化受到重视,在“退耕还草”“围封保护”等措施的开展下,部分草地退化区域形成了许多恢复演替群落。目前,关于高寒草甸恢复演替的研究已经有很多,且取得了大量的研究成果,可以在一定程度上解释植物群落演替的机制。王长庭等(2008)关于放牧干扰下高寒草甸的主要建群种和优势种更替的研究表明,牧压强度是影响高寒草甸植物群落物种多样性格局和演替的主要驱动力。曹广民等(2004)就土壤-牧草氮素供需状况变化对高寒草甸植被演替与草地退化的影响进行了研究,结果表明,土壤水分含量和氮素是高寒草甸演替发生的主要驱动因子。然而,目前关于高寒草甸不同演替阶段植物光合作用对环境适应机制的研究还很少。因此,本研究根据美国植物生理生态学家Bazzaz(1979)提出的比较“优势种生理生态特性”的方法,对甘南高寒草甸各演替阶段优势种主要光合生理特征的变化规律进行研究,试图回答以下问题:(1)验证Craig(1957)提出的中期物种丰富度学说(Mid-term species richness hypothesis);(2)不同演替阶段土壤含水量是否是决定植物光合性状差别的主导因子?通过对以上问题的研究进一步揭示植物种群在不同演替阶段的环境适应性对策,为进一步促进高寒草甸人工草场恢复年限的设定及草地资源的可持续利用提供科学依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于甘肃省甘南藏族自治州合作市附近(34.93°N,102.88°E),平均海拔3500 m左右,气候寒冷,太阳辐射强,属于温带大陆性气候。年平均气温2.0 ℃,0 ℃以上年积温1730 ℃,12—2月为最冷月,平均气温-8.90 ℃。年降水量约560 mm,6—8月降雨量最多,约占全年的75%;年平均霜期大于 270 d,属于典型的高寒草甸气候区。植被是典型的高寒草甸植物,以密丛型禾本科和莎草科植物为主,主要的优势种有矮嵩草(Kobresia humilis)、刺儿菜(Cirsium setosum)、三刺草(Aristida triseta)、蒲公英(Taraxacum mongolicum)、金露梅(Potentilla fruticosa)、珠牙蓼(Polygonum vivipurum)和鹅观草(Roegneria kamoji)等,主要的伴生种有披碱草(Elymus nutans)、火绒草(Leontopodium leontopodioides)、紫花苜蓿(Medicago sativa)、小米草(Euphorbia tatarica)和米口袋(Gueldenstaedtia verna)等。土壤为砂质壤土,呈微碱性,被归类为亚高山草甸土(杜国祯等,1995)。
1.2 试验设计
于2015年7月中旬到8月中旬在研究区选择5个典型群落样地(围封年限分别为 1、3、5、15、30 a),各阶段的优势种分别是草本植物刺儿菜(Cirsium setosum)、披碱草(Elmus dahuricus),紫苜蓿(Medicago sativa),半灌木植物沙蒿(Artemisia desertorum),以及灌木植物金露梅(Potentilla fruticosa)。每个样地中随机选取 8个0.5 m×0.5 m样方,调查物种多度、盖度、频度。
重要值(M)反映了某个物种在群落中的重要程度,以相对多度(RA)、相对盖度(RC)和相对频度(RF)的平均值来表示(岳明等,1997)。

丰富度指数(R)和Shannon-Wiener指数(H′)都是反映群落中物种多样性的指数(Ma et al.,1995;张金屯,2004),计算公式如下:

式中,S为物种数;pi为样方中第i种的个体数占所有种个体数的比例。
本研究区5个不同演替阶段高寒草甸植物群落变化特征如表1所示。
1.3 测定方法
1.3.1 环境因子的测定
使用多功能测温仪(型号DT-131)测量土壤表层(0~20 cm)温度(ts),每组重复3次。使用照度计(HT-8318)测量群落顶部(距离地面 60 cm处)光照度(Il),每组重复 3次。在每个样地内使用直径 5 cm的土钻沿着对角线采集 5钻土壤(0~20 cm),混合后立即装入铝盒,带回实验室称量湿重,然后在105 ℃下烘干称量干重,最后计算其土壤含水量(SWC),计算公式如下:

式中,msf为土壤湿重(g);msd为土壤干重(g)。
1.3.2 叶片光合性状的测定
2015年7月中旬到8月底,选择晴朗天气,于上午 9:00—11:00利用便携式光合仪(LI-6400,LI-COR,USA)测定各演替阶段优势种植物叶片的气体交换参数。测定时,选用开放气路,空气流速为 500 cm3·min-1,在外界 CO2浓度下大约为 350 μmol·mol-1条件下,用红蓝光源提供 1000 μmol·m-2·s-1光强进行测定,选择 3 株健康植物冠中部向阳枝条前端5~7片成熟完整的叶片进行测量,每个叶片记录3组数据用于统计分析。主要测定指标包括叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs),叶片瞬时水分利用率(WUEi)利用下式进行计算:

表1 不同演替阶段植被变化特征Table 1 Vegetation change characteristics at different successional stages

相对叶绿素含量采用 SPAD-502Plus手持式叶绿素测定仪测定,以测定净光合速率及蒸腾速率的植株为测定对象,每株选择5片叶片,在各个叶片中脉两侧均匀选取3个点,取平均值作为该叶片叶绿素含量,所测叶绿素含量是一个相对值。
将采集的叶片保存于湿润的封口袋中带回实验室,一部分直接称量叶片鲜质量,然后在 75 ℃烘箱中烘48 h,称量,计算每个叶片含水量;另一部分用超纯水洗净并自然晾干,于80 ℃下烘干48 h至恒重,而后在过量O2中完全燃烧并收集CO2,然后用气体质谱仪Delta Plus测定试样中的δ13C含量,测量精度小于 0.1‰(Craig,1957)。计算公式如下:

式中,mlf为叶片湿重(g);mld为叶片干质量(g);Rsam为植物样品的ω(13C)/ω(12C)值;Rsta为通用的标准化石标样的ω(13C)/ω(12C)。
1.4 数据处理
运用Microsoft Excel 2007对数据进行整理与计算;运用Canoco 4.5软件绘制物种-样地的PCA二维排序图和环境-植物性状的RDA二维排序图;运用SPSS 19.0软件中的单因素方差分析(One-way ANOVA)对不同演替阶段优势叶片各指标进行差异显著性分析;运用Origin 9.0软件作图。
2 结果与分析
2.1 不同演替阶段植物群落组成
排序可以实现群落数量分类,本研究采用PCA排序法对群落演替进行定量分析,分析结果见表 2和图1。
主成分分析是通过原有变量线性组合出新变量(即主成分)来反映事物发展的主要方向,该分析中4个轴上的特征值分别为0.363、0.169、0.124、0.082(表2),4个成分对样方数据的总累积贡献量为73.8%。前两个轴对物种变量的解释量最大,分别为36.3%和16.9%,可以作为主成分轴。
由图1可知,演替第1、3、5年群落样方基本位于第三象限,样方距离比较近。演替第 15年群落样方位于第四象限,样方间差异性较显著。演替第 30年群落样方位于第一和第二象限之间,各阶段在排序图上形成3个明显的样方聚居区。带箭头的射线代表物种,样方圆投影在射线上的点到箭头的距离可反映该样方与物种的关系,沿着箭头方向的为增大,反之减小,以此反映相应样方中该物种多度的大小,即处于某个物种射线方向上的样方该物种数量较多,处于反方向上的样方该物种数量较少,而处于原点处的样方内该物种的多度值大致等于其平均值。由图1可知,演替1、3、5年(依据围封)群落中优势种分别为刺儿菜、披碱草和紫花苜蓿,且群落中主要物种数量都较少,演替第 15年样地内群落物种丰富度(36.63)和多样性指数(4.24)都明显升高,优势种为半灌木植物沙蒿,主要伴生种有球花蓼(Polygonum glomerata)、羊茅(Festuca ovina)、大戟(Euphorbia pekinensis)、龙胆(Gentiana scabra Bunge)等。演替30年达到顶级群落,优势种为灌丛植物金露梅(重要值为0.327),该群落内主要伴生种为鹅绒委陵菜(Potentillaanserinal)和野草莓(Fragaria vesca),物种数与前一演替阶段相比较少。结合表1和图1,发现整个演替过程形成了草本→半灌木→灌木的演替序列,而前3个草本植物群落按功能型又可以看成是一个由杂生草群落→禾本科群落→豆科群落的演替序列。
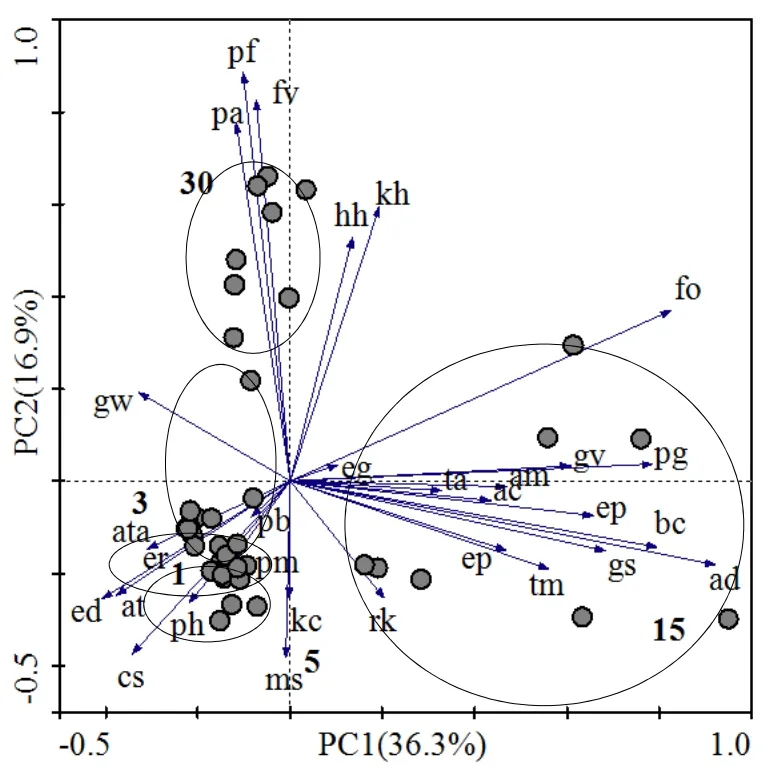
图1 不同演替阶段主要物种-样方PCA二维排序图Fig. 1 The two-dimensional sequence of PCA in the different successional stages1,3,5,15,30分别代表演替年限;cs,刺儿菜;at,甘青蒿;gw,老鹳草;ata,细叶亚菊;ph,毛连菜;er, 节节草;ed,披碱草;ms,紫花苜蓿;pb,二裂委陵菜;pm,多裂委陵菜;pa,鹅绒委陵菜;rk,鹅观草;kc,恰草;ad,沙蒿;gs,龙胆;ep,小米草;tm,蒲公英;ep,大戟;eg,火绒草;ta,唐松草;ac,银莲花;am,黄芪;ar,小花草玉梅;gv,米口袋;pg,球花蓼;fo,羊茅;ch,柴胡;kh,矮蒿草;hh,狗娃花;fv,野草莓;pf,金露梅1, 3, 5, 15, 30 respectively represent the successional stage. cs,Cirsium setosum; at, Artemisia tangutica; gw, Geranium wilfordii; ata,Ajania tenuifolia; ph, Picris hieracioides; er, Equisetum ramosissimum; ed,Elymus dahuricus; ms, Medicago sativa; pb, Potentilla bifurca; pm.,Potentilla multifida; pa, Potentilla anserinal; rk, Roegneria kamoji; kc,Koeleria cristata; ad, Artemisia desertorum; gs, Gentiana scabra Bunge;ep, Euphrase pectinata Ten; tm, Taraxacum mongolicum; ep, Euphorbia pekinensis; eg, Echinops gmelini; ta, Thalictrum aquilegifolium; ac,Anemone cathayensis Kitag; am, astragalus membranaceus; ar, Anemone rivularis Buch; gv, Gueldenstaedtia verna; pg, Polygonum glomerata; fo,Festuca ovina; bc, Bupleurum chinense; kh, Kobresia humilis; hh,Heteropappus hispidus; fv, Fragaria vesca; pf, Potentilla fruticosa

表2 样方数据PCA分析4个轴上的特征值Table 2 Quadratic Data PCA analyzes eigenvalues on four axes
2.2 不同演替阶段环境因子的变化
从演替第1年到演替30年,环境因子均发生了变化。如图2所示,在演替的前3个时期,土壤含水量一直处于缓慢增长的趋势,从演替的第 15年起,SWC上升幅度逐渐变大,演替至第30年达到最大值0.390 g·g-1(图2A);ts在演替第1年最高,而后逐渐下降,在演替第 15年达到最低,为18.622 ℃,从演替第15年到30年以上又开始呈上升趋势(图2B);由图2C可以看出,Il在演替的第1~3年变化不明显,第3年之后,开始呈明显下降趋势,并且各阶段差异显著(P<0.05)。
2.3 不同演替阶段植物叶片光合性状的变化
本研究对不同演替阶段群落内优势种的叶片光合性状进行加权计算,得到叶片光合性状平均值。如图3所示,不同演替阶段优势种叶片δ13C值随演替的进行呈规律性下降趋势。C3植物的 δ13C值变化范围在–35‰~-20‰之间,说明甘南高寒草甸植物都属于C3植物(图3A),各阶段C3植物叶片δ13C值差异显著(P<0.05)。从演替前期到后期,叶片净光合速率和瞬时水分利用效率变化趋势大致相同,均呈下降-上升-下降趋势,在演替第 5年达到峰值,分别为 16.987 µmol·m-2·s-1、3.762µmol·mmol-1(图3B,图3D)。随演替进程,气孔导度显著升高(P<0.05),而蒸腾速率则呈下降趋势(图3C,图3E)。由图3F可知,叶片含水量随演替的进行呈先增大后减小的趋势,演替第 15年达到5.207 g·g-1,显著高于其他各阶段(P<0.05)。由图3G可知,叶片相对叶绿素除演替第1年较高外,其他演替阶段间无显著差异(P>0.05)。
2.4 不同演替阶段叶片光合性状与环境因子的RDA分析
以甘南高寒草甸不同演替阶段5个优势种植物叶片光合性状指标代表物种(species),以3项环境因子作为环境因素(environments),对 2组变量进行RDA分析,得到排序结果(表3,图4)。结果表明,LWC、Gs与 SWC呈正相关,相关性LWC>Gs;SPAD、Tr、Pn、WUEi、δ13C 与 SWC 呈负相关,相关性大小排列为 WUEi>δ13C>Tr>Pn>SPAD。ts、Il与 SPAD、Tr、Pn、WUEi、δ13C 呈正相关,与LWC、Gs呈负相关。由表3可知,土壤含水量 SWC与第 1排序轴存在显著正相关(r=0.871),土壤温度ts、光照度Il与第1排序轴呈显著负相关(r=-0.790,r=-0.891),这 3个环境因子与第2排序轴的相关性均不显著。

图2 不同演替阶段环境因子的变化Fig. 2 Changes of environmental factors in the different successional stages

表3 环境因子与排序轴之间的相关性Table 3 Correlations between environmental factors and ordination axes

图3 不同演替阶段优势种光合性状的变化Fig. 3 Changes of photosynthetic traits in the different successional stages
3 讨论
3.1 演替梯度上群落结构的变化

图4 不同演替阶段叶片光和形状与环境因子的RDA分析Fig. 4 RDA analysis of leaf Photosynthetic traits and environmental factors in different succession stages
群落的演替过程主要是通过优势种群的消亡与增加来体现的。在漫长的演替过程中,群落物种组成、盖度、高度、密度等都会发生改变(张梦弢等,2015)。关于群落演替过程中物种丰富度的变化格局,主要有两种假说——经典学说和中期物种丰富度假说(Baniya et al.,2009;Bazzaz,1996)。大多数研究表明,演替初级阶段,群落物种数量、种类极少,随着演替的进行,物种数量、种类开始增加,直到达到顶级群落时多样性最大、物种数最多(夏艳菊等,2018),与经典学说一致。然而,本研究与前人研究结果略有不同,在演替初始阶段,样地内主要以一年生草本植物居多,群落结构简单,仅存在少量伴生物种,如老鹳草(Geranium wilfordii)、甘青蒿(Artemisia tangutica)等;随着时间的推移,尤其是在演替第 15年,群落物种丰富度、复杂度,植被类型等都发生了明显的改变,群落结构更趋复杂化,多样性显著增加,优势种作用越来越重要;演替到 30年以上时,金露梅占据主导优势地位,多样性又趋于降低,这一现象符合中期物种丰富度假说,即演替进程中的物种丰富度呈单峰模型,演替中期物种丰富度最高。这是因为群落演替至中期时,垂直和水平的异质性最大,导致群落物种多样性达到最大,而当演替至后期时,优势度很强的物种出现或优势种存在化学他感作用,使群落物种多样性逐渐降低(王世雄等,2010)。
3.2 演替梯度上光合性状的变化
光合作用是植物进行生理和生命活动的基础,与植物生长、发育有密切的关系。本研究对高寒草甸不同演替阶段 5个优势种的光合性状分析显示(图 3),这些物种的叶片 δ13C、Pn、Tr、WUEi、Gs、LWC及SPAD在不同演替阶段存在明显差异。演替进程中,植物叶片气孔的闭合程度会直接影响叶片的δ13C值,进而影响WUEi。本研究中,随着演替的进行,各阶段优势植物δ13C呈现一定的规律性,前期优势种刺儿菜叶片δ13C明显高于后期优势种金露梅的叶片δ13C,即草本植物的WUEi高于灌丛植物,这可能是因为草本植物的水分传输系统和传输路径强于木本植物,也可能是因为演替前期植物适应旱生环境的能力更强(吴统贵,2009)。然而,Llambi et al.(2003)研究表明,随着演替的进行,优势种的水分利用率不断增加,与前期优势种相比,后期物种可以通过降低气孔导度来减少水分损失,从而有利于自身立足于不利环境中。本研究中,以刺儿菜、披碱草及紫花苜蓿为优势种的早期草本群落的 Pn大于以金露梅为优势种的后期灌丛群落,陈世伟(2015)研究也表明,演替早期的植物都具有较高的Pn、Tr,随着演替的进行,各指标逐渐下降。这是因为演替前期群落为草本植物群落,后期为木本植物群落,演替的早期生境是充满阳光的开放环境,很多演替早期的植物具有阳生植物的特性,后期植物具有阴生植物的特性,阳生植物的Pn较阴生植物稍低,故演替前期植物的Pn大于后期植物(李庆康,2002)。安慧等(2007)曾对黄土高原植被不同演替阶段优势种的蒸腾速率进行对比,发现草本植物群落蒸腾速率大于灌木群落,吴统贵(2009)研究也指出演替后期优势种的蒸腾速率低于演替早期。本研究中,演替 30年以上优势种Tr高于演替第1年,这可能是由于高寒草甸特殊的地理环境,但Tr整体符合前期高于后期的结论。叶绿素也与光合作用密切相关,其含量和比例是植物适应和利用环境因子的重要指标。本研究中,演替第1年叶绿素含量明显高于其他阶段,后保持在一个相对稳定状态。
3.3 不同演替阶段光合性状对环境因子的响应
植物光合作用不仅取决于植物本身的遗传特性,同时还受外界环境条件的影响和制约(吴统贵等,2008;冯晓钰等,2018)。演替顶级理论(Climax)认为任何一个具体的地区,一般的群落顶点取决于该地区的气候特征(Clement,1916)。土壤、气候等环境因子各有其存在的形式和规律,并且以各种组合方式影响植物的生长、发育、分布及其多样性,是影响植被群落演替、分布的外在因子。以往研究表明,土壤含水量是推动演替进行的主要环境因子(韦兰英等,2006),而本研究中RDA分析结果表明(图 4),演替过程中环境因子对优势种叶片光合性状影响强度大小为:Il>SWC>ts,即光照是演替进程中影响叶片光合性状的主导因子,土壤含水量次之,这与演替不同阶段入射到植被群落上光强和光质的差异有关(管铭等,2014)。在高寒草甸,植被群落演替前期,恶劣的环境条件会导致物种生存难,盖度小,植物处于开放的阳生环境中,Il及ts都较高,此环境更适宜阳生植物生长,具有最大Pn的刺儿菜、披碱草在此环境中成功生存,同时较高的Tr可以显著降低植物叶片温度,从而保护前期植物免受高温胁迫;而到了演替后期,群落环境明显好转,植物种类显著增加,盖度增大,且出现灌丛植物,群落下层 Il减少,ts在上层植物的保护下明显优化。这进一步说明不同演替阶段光环境及快速变化的光照是影响植物生长、生存,进而促进物种更新与群落演替的主导因子。除了光照度,植物光合性状还与土壤含水量有关,土壤水分过高或过低均会降低植物叶片的Pn、WUE、Gs(马剑英等,2008;王亚婷等,2017)。一般而言,土壤水分亏缺将引起气孔和非气孔因素限制间接导致光合指标产生差异。本研究中,随着演替的进行,优势种叶片Gs和LWC随SWC的增加而升高,δ13C、Pn、Tr、WUEi等随SWC的增加而降低,与前人研究结果相似,这是因为当土壤水分的可利用性降低时,植物为减少体内水分散失往往关闭部分气孔,造成叶内CO2分压降低,Ci/Ca减小,从而使光合产物的δ13C 值增大,WUEi提高(刘旻霞等,2016;Farquhar et al.,1982)。这说明植物水文过程也能够影响群落演替过程。
4 结论
甘南高寒草甸群落演替经历了草本-半灌木-灌木的演替过程,物种的丰富度和多样性指数均出现了较大波动,从前期到后期呈现先增加后降低的变化趋势;从演替前期到后期,叶片光合性状中,除叶片SPAD外,其他性状均随演替进程呈显著差异(P<0.05);环境因子中,土壤含水量(SWC)随演替进行呈增大趋势,土壤温度(ts)和光照度(Il)则呈降低趋势,其中,光照度是该区演替发生及影响光合性状的主导因子。相比于演替前期,后期优势种叶片光合性状的可塑性更强,对环境因子的变化适应性更高,演替过程呈现由有机物质积累较强转变为有机物保持能力较强的变化。
