喀斯特森林宜昌润楠蒸腾耗水规律及其与环境因子的关系
2019-01-04赵文君舒德远李成龙崔迎春刘延惠侯贻菊丁访军
赵文君,舒德远,李成龙,崔迎春,刘延惠,吴 鹏,侯贻菊,丁访军
(贵州省林业科学研究院,贵州 贵阳 550005)
蒸腾在岩石—土壤—植物—大气连续体(SPAC)水热传输过程中占有极为重要的地位[1],树干液流作为衡量植物蒸腾耗水的一个重要生理指标,能够反映植物生理特性和环境因素对植物水分利用的综合调节作用[2]。热脉冲法、组织平衡法和热扩散式探针法[3-4]实现了对树干液流的连续监测,可推测植株或整个林分蒸腾量,对理解树木的水分生理过程、水分平衡及植物生理生态行为与森林生态系统功能稳定性的关系有重要意义[5-8]。
喀斯特森林是在可溶性碳酸盐发育的喀斯特地貌上生育的森林,其生境与相同气候条件下的常态地貌上常绿阔叶林有明显的不同[9]。喀斯特森林土层浅薄且不连续,贮水能力低且岩石渗漏性强,普遍存在不同程度的水分亏缺,生态水文过程较为复杂,形成了喀斯特森林树种不同于其他地区树种的水分生理特性和水分利用策略。而对喀斯特森林树干液流相关研究较少,仅对青冈栎Cyclobalanopsis glauca[10]、短萼海桐Pittosporum brevicalyx[11]、细 叶 青 冈Cyclobalanopsis gracilis[12]、天峨槭Acer wangchii[13]等树种有过研究报道。目前对喀斯特森林树木蒸腾及生理生态过程仍缺乏足够认识。为此,本研究以喀斯特森林顶极群落亚优势种宜昌润楠Machilus ichangensis为研究对象,利用热扩散探针技术对其树干液流进行连续监测,并结合气象和土壤因子的同步监测,分析其液流速率变化、蒸腾日变化和季节变化及与环境因子的响应关系,以期深入了解喀斯特森林生境下树种的蒸腾规律及生态适应性,为退化的生态系统恢复和重建中树种选择提供依据,为土壤水分植被承载力评价和生态环境建设、管理提供理论依据。
1 研究区概况和研究方法
1.1 研究区概况
研究在茂兰国家级自然保护区内(107°52.2′~108°5.7′E,25°9.3′~ 25°20.8′N)贵州喀斯特森林生态系统定位观测研究站开展。该区属中亚热带季风湿润气候,年平均气温15.3 ℃,≥10℃积温4 598.6℃;年平均降水量为1 752.5 mm,集中分布于4ü10月;年平均相对湿度为83.0%;无霜期为315 d;全年日照时数为1 272.8 h,日照百分率为29.0%;太阳年辐射总量为63 289.80 kW/m2,海拔在430.0~1 078.6 m之间。该区属于典型的喀斯特生境,裸岩率在80%以上,土壤稀少,土层浅薄,土壤为石灰土。该区是目前世界上同纬度地区残存下来仅有的、原生性强且相对稳定的喀斯特森林生态系统,同时也是喀斯特区原生性森林分布面积最大的地区[9,14]。
对茂兰喀斯特峰丛中部常绿落叶阔叶混交林中4个30 mh30 m固定样地进行了调查,乔木层郁闭度为0.75,主林层高11~25 m,次林层高6.5~9.0 m,研究区优势树种主要有云贵鹅耳枥 Carpinus pubescens、圆果化香Platycarya longipes、朴树 Celtis tetrandra、黄连木 Pistacia chinensis等落叶树种,天鹅槭Acer wangchii、青冈栎 Cyclobalanopsis glauca、细叶青冈Cyclobalanopsis gracilis、腺叶山矾 Symplocos adenopus等常绿树种。亚优势种主要有宜昌润楠Machilus ichangensis、裂果卫矛Euonymus dielsianus等常绿树种。林下灌木树种主要有南天竹Nandina domestica、长梗罗伞 Brassaiopsis glomerulata var.longipedicellata、九里香 Murraya exotica、中华野独活 Miliusa sinensis等,灌丛平均高度为1.9 m,盖度为20%。草本植物主要有麦冬Ophiopogon japonicus、马蓝 Baphicacanthus cusia、冷水花 Pilea notata等,草本盖度为10%[11]。
1.2 树干液流的测定
观测样地位于峰丛中部较平缓处,坡度13°,坡向为西向,海拔为700~730 m。宜昌润楠是该林分的亚优势树种之一,其重要值为2.73[15]。根据样地调查结果,选取树干通直、无病虫害、不偏冠、生长良好、径级适中的3株宜昌润楠为样树(特征见表1),采用热扩散探针法于2015年7月至2016年8月对宜昌润楠树干液流进行连续测定,将TDP探针安装在树干北侧胸高1.3 m处,为防止雨水流入和太阳辐射引起温度波动,在外面包裹防辐射薄膜,探针另一端与数据采集器CR1000连接,以10 min时间间隔自动采集数据。
根据Granier经验公式[16]来换算树干液流密度(Js):
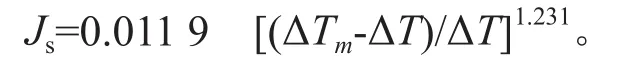
式中:Js为液流密度(瞬时液流速率)(g/m2·s),即单位时间通过单位边材面积的液流量[23];ΔTm为上下探针间的最大昼夜温差(℃);ΔT为瞬时温差(℃)。
小时蒸腾量通过下式计算:
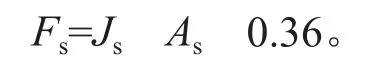
式中:Fs为小时蒸腾量(g/h);As为胸径处边材面积(cm2);0.36是时间换算系数[23]。在Fs基础上通过累加便可以计算得到单日蒸腾量Fd(kg/d)。
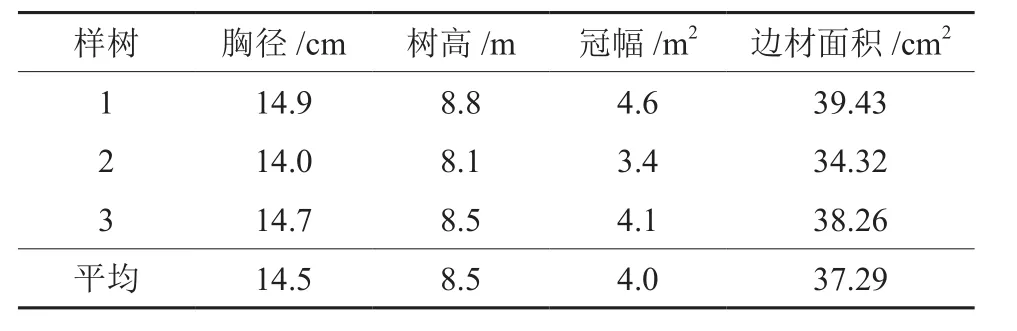
表1 宜昌润楠样树基本特征Table 1 Basic characteristic of sample trees of M.ichangensis
1.3 环境因子的测定
环境因子由安装在观测塔上的Campbell自动气象站测定,数据采集时间与树干液流同步,主要包括:太阳辐射(R,x1)、气温(Ta,x2)、相对湿度(RH,x3)、风速(WS,x4)、降水量(P,x5)、不同层次的土壤含水量(SWC10,x6;SWC20,x7;SWC40,x8)等。
1.4 数据处理
采用Excel2007进行数据处理,SPSS16.0进行方差分析、相关性分析、逐步回归分析及通径分析等;通径系数计算方法参考文献[17]。用Excel2007、origin9.1对液流速率的日变化、蒸腾量季节变化及其环境因子关系进行作图。
2 结果与分析
2.1 宜昌润楠树干液流速率日变化规律
不同季节各选取典型晴天3日数据分析液流速率的日变化(见图1),不同季节宜昌润楠液流速率呈现明显的昼高夜低变化,白天为单峰曲线,夜间为平缓的波动曲线。白天,树干液流用于蒸腾,将其定义为日间蒸腾阶段。夜间,液流用于补充日间的树体水分消耗,定义为夜间补水阶段。春、夏、秋、冬各季日间树干液流速率平均值分别是夜间的8.95倍、14.32倍、10.0倍、5.24倍。方差分析表明,树干液流昼夜差异显著(P<0.05)。

图1 不同季节树干液流速率和太阳辐射日变化过程Fig.1 Diurnal variation of sap flow velocity and solar radiation in different seasons
各季节树干液流随太阳辐射而变化,但启动和结束时间相对于太阳辐射存在一定的时滞性(见图1)。液流于6:30ü8:30间启动,而后随着气温和太阳辐射增加出现快速上升阶段,在11:00ü12:00达到高值;夏季能维持约4 h,约14:00出现峰值;秋季能维持约2 h,12:30出现峰值;春季、冬季介于两者之间,约13:00出现峰值。随着气温及太阳辐射减弱其液流下降,春夏季在17:00(秋冬季在16:00)开始迅速下降,春夏季在20:00ü21:00(秋冬季在19:00ü20:00)达到低值,并维持低水平波动,且前半夜(20:00ü23:00)高于后半夜(0:00ü 5:00)(液流值趋于0),直至第二日液流启动。由冬季到夏季,液流启动时间逐渐提前,而到达峰值时间及降到低值的时间逐渐推后,日间树干液流历时从冬季11.5 h到夏季可延长达15 h,可能与太阳辐射时间的长短有关。整个观测期间,宜昌润楠日均液流速率为17.83 g·m-2s-1,范围为0.06~95.14 g·m-2s-1,各季节树干日均液流速率(g·m-2s-1)表现为夏季(25.00f0.81)>秋季(20.09f1.03)>春季(11.91f0.62)>冬季(7.00f0.30),且各季节之间差异显著(P<0.05)。
2.2 不同典型天气下宜昌润楠树干液流速率变化规律
图2为2015年8月典型天气(晴、阴、雨)条件下宜昌润楠树干液流的变化。由图2可以看出,晴天、阴天液流的日变化相似,晴天为尖峰曲线,阴天变化较平缓。晴天液流启动约在7:30,滞后太阳辐射约1 h,13:00达到峰值,为291.57 g·m-2s-1,21:00降到低值,整个日间蒸腾持续13.5 h。阴天液流启动较晚,约在8:10,滞后于太阳辐射约2 h,14:30 达到峰值,为 143.15 g·m-2s-1,19:00 降到低值,日间蒸腾持续时间较短,仅10.5 h。雨天树干液流值明显低于晴天和阴天,12:50达到峰值,为59.10 g·m-2s-1,呈现上下波动变化,无明显规律,滞后于太阳辐射约2 h,降水量、降水历时、降水强度等均会影响液流速率的变化[18]。
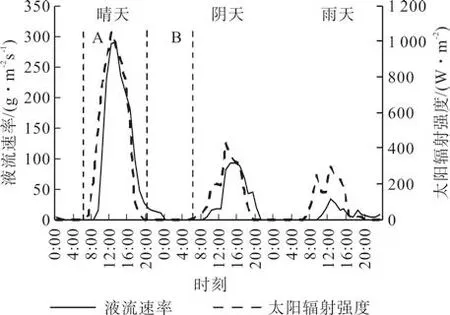
图2 不同典型天气条件下树干液流速率及太阳辐射日变化Fig.2 Diurnal variation of sap flow velocity and solar radiation under different weather conditions
观测期间不同天气条件下液流速率(g·m-2s-1)日均值分别为晴天(41.93f1.43)>阴天(12.73f 0.63)>雨天(8.82f0.29)。各天气条件下日间时段(A)树干液流速率平均值(g·m-2s-1)分别为46.44、21.00 、13.52,夜间时段(B)树干液流速率(g·m-2s-1)较接近,分别为 1.52、1.68 、1.73。不同天气条件下,昼夜贡献率分别为96.82%、3.18%,92.58%、7.42%和88.61%、11.39%,夜间贡献率为雨天>阴天>晴天。不同天气条件间、昼夜间差异显著(P<0.05)。
2.3 宜昌润楠蒸腾季节变化规律
宜昌润楠日均蒸腾量与气温、太阳辐射同步变化,表现出年内及季节间差异(见图3)。整树日蒸腾量平均为(5.75f0.31)kg/d;11月—次年2月蒸腾量较低,1月最低为1.54 kg/d; 3月—6月日蒸腾量开始增加,达到4.00 kg/d以上; 7月—10月蒸腾较大,7月达到峰值(12.42 kg/d)。宜昌润楠日均蒸腾量表现出季节间差异显著(P<0.05),各季日蒸腾量(kg/d)大小依次为夏季(7.57f 0.57)>秋季(6.75f0.72)>春季(4.00f0.54)>冬季(2.35f0.20)。
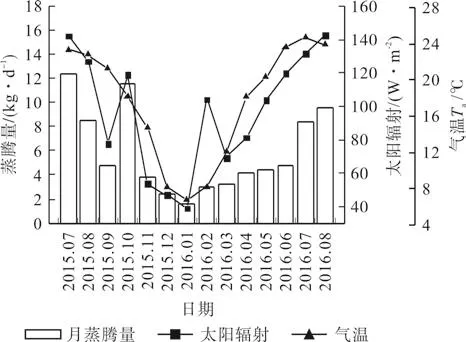
图3 宜昌润楠月均日蒸腾量与环境因子的月均值变化Fig.3 Monthly mean variation among daily transpiration in M.ichangensis and environmental factors
2.4 蒸腾与环境因子的关系
不同天气条件、不同时间尺度下影响蒸腾的环境因子及其影响方式、作用大小各不相同,且各环境因子之间存在相关关系,因此进一步对各环境因子与蒸腾进行通径分析,探究各环境因子的总影响、直接影响和间接影响。
2.4.1 小时尺度下蒸腾与环境因子的关系
表2可以看出,不同天气条件下,影响全天及白天蒸腾的主要环境因子是R、RH、T,影响夜间蒸腾的环境因子晴天主要是SWC40,阴天主要是Ws和RH,雨天主要是P。从逐步回归中各因子进入的顺序判断,小时尺度上各因子对蒸腾影响最大的是R,最小的是P、SWC。从影响方式来看,RH在各种天气各时段与蒸腾均呈显著负相关;R、Ws在全天和白天与蒸腾呈显著正相关;Ta除在阴天夜间与蒸腾呈显著负相关外,其余各天气时段下均呈显著正相关;P在雨天与蒸腾呈显著的正相关。SWC除在雨天夜间与蒸腾呈正相关外,其余各天气时段均呈显著负相关。从环境因子对蒸腾直接、间接影响看,在全天和白天,直接影响主要表现为RH>R>T(除阴天全天表现为R>RH>T);间接影响,晴天、阴天主要表现为Ws>R>RH,雨天为R>RH>SWC10>T(全天)和R>SWC10>T>RH(白天)。可见,RH、R(除雨天外)主要通过直接作用影响蒸腾,Ws(晴天、阴天)及R(雨天)主要通过对其他因子影响而间接影响蒸腾。
在通径分析的基础上,以各环境因子为自变量,蒸腾量为因变量,进行逐步回归分析,得到最优模型(见表3)。从全天来看,不同天气条件下,晴天和雨天各环境因子对蒸腾的解释程度相当在70%以上,高于阴天的61.8%。从昼夜来看,白天各环境因子对蒸腾的解释程度显著高于夜间。
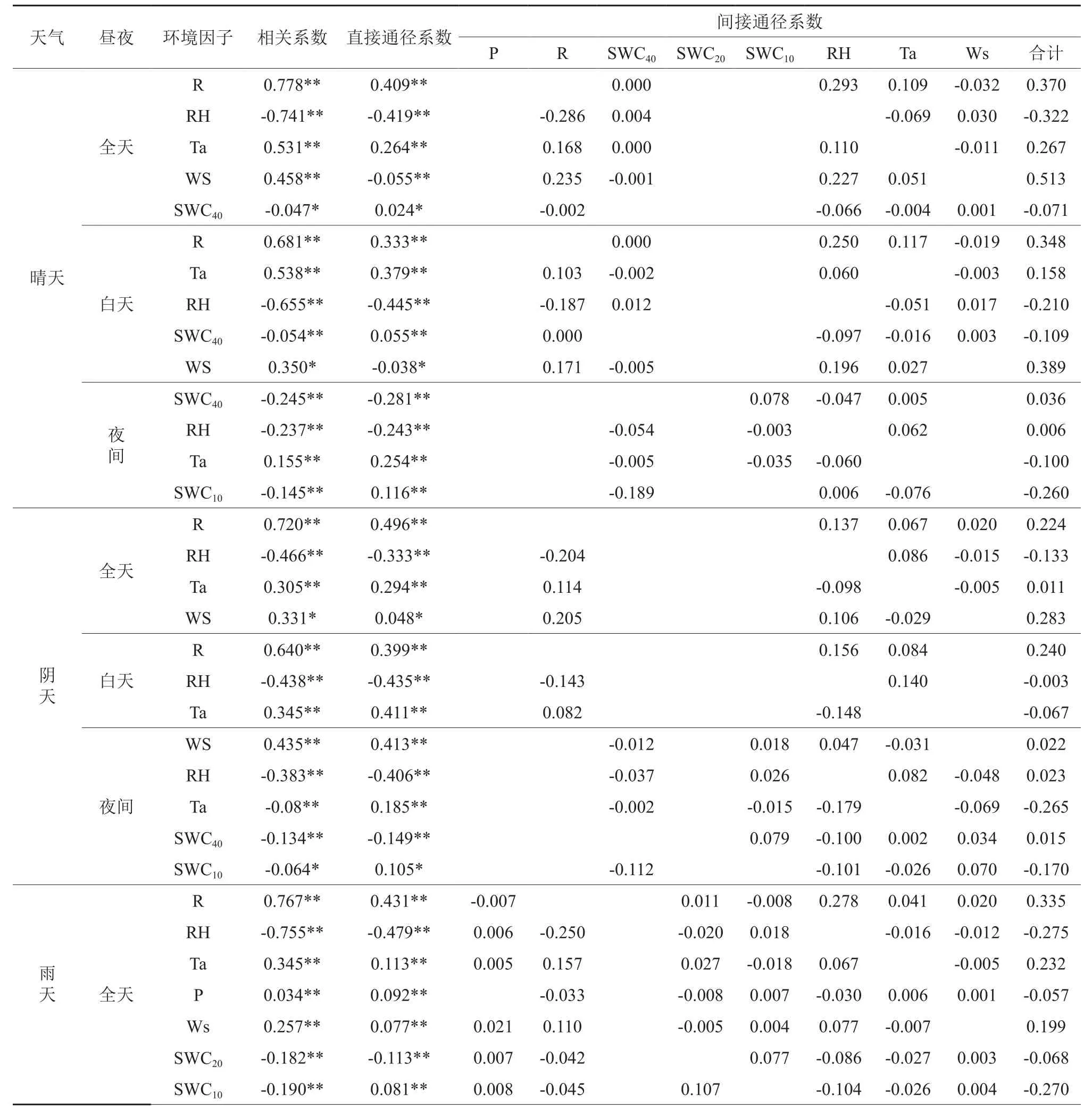
表2 小时尺度下环境因子对蒸腾昼夜变化的通径分析结果†Table 2 Path analysis results among diurnal variations of transpiration and environmental factors on the hourly scale
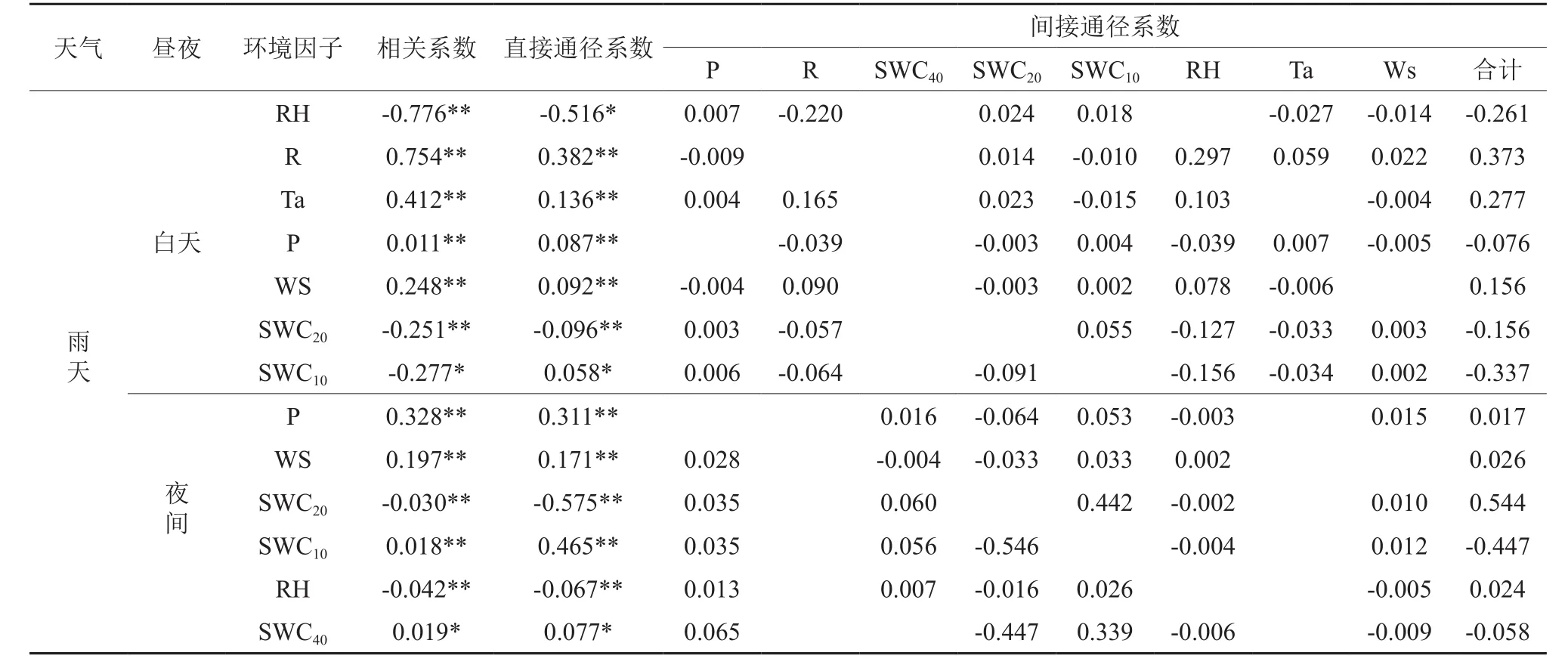
续表2Continuation of table 2

表3 蒸腾量昼夜变化与环境因子逐步回归分析结果Table 3 Stepwise regression results among diurnal variations of transpiration and environmental factors
2.4.2 日尺度、月尺度下蒸腾与环境因子的关系
表4可知,在不同的尺度下,影响蒸腾的主导因子会发生变化,日尺度下,春、夏、冬季太阳辐射、相对湿度是影响植物蒸腾的主要因素;秋季除太阳辐射影响外,SWC10也是影响蒸腾的主要因素。但任何情况下太阳辐射都是影响蒸腾的主要环境因子,太阳辐射的强弱决定气孔开张程度,又决定空气温度和相对湿度的变化,从而直接影响蒸腾作用[19]。各回归方程及回归系数都达到了显著水平,日尺度下各季节环境因子对蒸腾量解释程度除冬季较低(63.8%)外,其余季节都达到了85%以上;月尺度下全年环境因子对蒸腾量解释程度达到了78.9%。
3 讨论与结论
与多数树种一致[19-22],晴天宜昌润楠树干液流速率日变化呈单峰曲线,是因为白天树木蒸腾速率大,木质部水柱产生张力,大量的水分涌入根部,破坏了根部的渗透势梯度以被动方式吸入体内,而夜间的液流主要是由根压引起,根系以主动方式吸水,补充白天植物蒸腾损失的水分,恢复植物体内的水分平衡[21]。任启文等[23]研究冀北华北落叶松得出,晴天液流速率变化呈宽峰型,10:00ü16:00保持较高水平,而本研究呈窄峰型,12:00ü14:00保持较高峰值,也有一些植物因蒸腾午休或干旱胁迫而呈现双峰型[24-25]。可见,树干液流速率日变化与植物种类、立地条件、土壤供水、气象条件等密切相关。
不同天气条件下,树干液流速率的日变化表现为晴天>阴天>雨天,这与天峨槭[13]、缙云山典型树种[18]研究结果一致。晴天通常伴随着高温和强辐射,促进叶片气孔开放,减少气孔阻力,植物蒸腾活动强烈,增强液流活动[26]。阴雨天受降雨事件及光照的间断性的影响,温度、太阳辐射降低和空气湿度的增高,抑制了叶片气孔内外蒸汽压梯度的增加,植物蒸腾活动减弱[27-28]。 夜间树干液流贡献率雨天高于晴天、阴天,雨天蒸腾减弱,夜间因土壤水分条件的改善,树干能充分吸水,雨天夜间液流速率与日间差距较晴、阴天小,而使夜间树干液流贡献率雨天高于晴天、阴天。张璇等[18]发现导致雨天夜间液流率大的原因是降雨增加了土壤含水量。胡兴波等[29]发现白榆的夜间补水量与日降水量呈正相关。这与池波等[30]对兴安落叶松的研究不一致,夜间贡献率为晴天>雨天>阴天,这可能与所处地区立地条件、降水事件等不同有关。
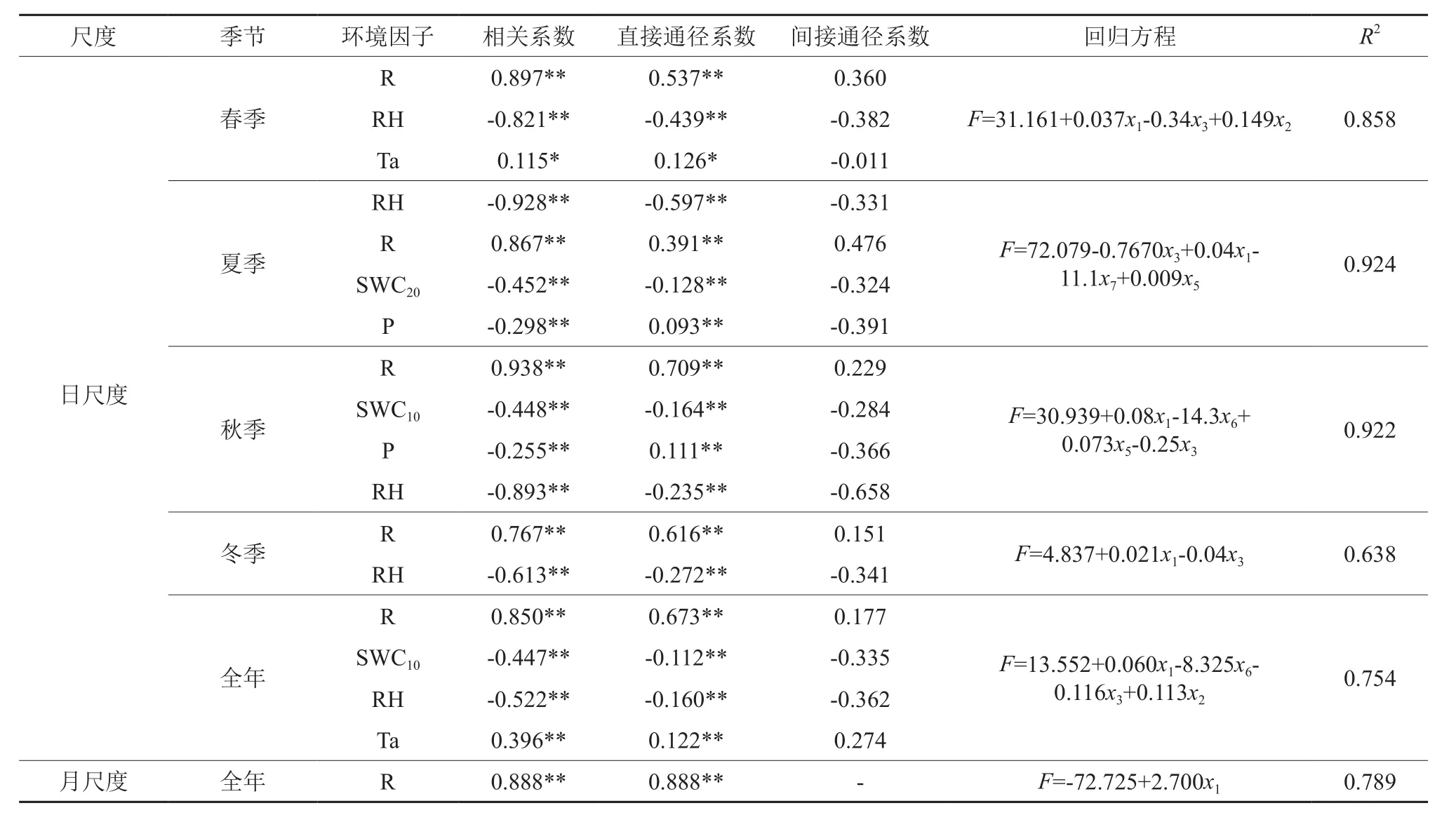
表4 日和月尺度下蒸腾与环境因子的逐步回归分析结果Table 4 Stepwise regression results among transpiration and environmental factors on daily and monthly scales
宜昌润楠日均蒸腾量为(5.75f0.31)kg/d,其值高于同处喀斯特森林的海桐(2.10 kg/d)[11]、细叶青冈(3.37 kg/d)[12]、天峨槭(5.08 kg/d)[13]。同一环境下树种蒸腾量之间的差异主要是由胸径、夜流速率不同所致,也与树木的生理结构、空间分布结构、树形特征、生长期等因素有关[18]。与同处亚热带气候区的其他树种相比,远低于岩溶地区的青冈栎(30~80 kg/d)[10]、荷木干季(23.88f12.52) kg/d和湿季(32.48f19.92) kg/d的平均蒸腾量[31]。这可能是宜昌润楠长期适应喀斯特多石少土生境形成的低耗水水分利用策略,且各样树立地条件、气候条件、土壤供水以及土壤肥力等因子的不同,均会导致植物对水分的利用产生差异[20]。宜昌润楠蒸腾量年内变化较大,11月至翌年2月属于低谷期,3ü6月属于上升阶段,7ü10月处于高峰期,季节差异明显,表现为夏季>秋季>春季>冬季。青冈栎[10]、细叶青冈[12]、兴安落叶松[30]都有相同的结论,这主要是与树木生长发育的特性有关,在生长旺盛的季节液流量大。
环境因子对树干液流的影响的结论存在差异,赵仲辉等[19]研究表明影响湖南会同杉木瞬时液流速率的主要是太阳辐射、空气温度,影响日液流的主要是空气温度、土壤含水量、太阳辐射,影响月液流的主要是冠上气温和相对湿度。丁访军等[32]对毛竹研究表明非生长季树干液流速率与空气温度的相关性最强,而生长季节与光合有效辐射的相关性最强。徐先英等[33]研究表明整个生长季,VDP或T对梭梭起主导作用,而太阳辐射对怪柳和白刺起主导作用。本研究随着时间尺度的增大,影响树干液流的主导因子也随着减少,只有太阳辐射在任何情况下都是影响液流的主导因子,其他环境因子则对某些观测时段的液流产生作用。各研究都反映出液流速率对环境因子的响应存在时间尺度、昼夜间、天气间、季节间的差异。宜昌润楠表现为白天高于夜间,晴天、雨天高于阴天,夏、秋、春季高于冬季。
喀斯特地区生境异质性大,加之多石少土的环境,使得喀斯特地区树木水分生理生态机制有其独特性,大量像宜昌润楠这样适应岩溶环境的植物往往能增加降水在表层岩溶带的停留数量及停留时间,并能吸收利用这部分水分,增加生态系统的有效水资源[10]。有研究表明细叶青冈[12]、青冈栎[10]在干旱季节日蒸腾量一直维持较高的水平,并未随土壤含水量的减少而下降,其水分来源可能很大程度依赖于表层岩溶带或其他渠道。对喀斯特地区林木的水分生理生态机制还需结合小生境类型及植物的生态适应机制等因素进一步深入研究。
