肉牛及奶牛生长阶段饲粮的氨基酸限制性顺序及理想氨基酸模式研究现状
2019-01-03刁其玉
■李 媛 刁其玉 屠 焰
(中国农业科学院饲料研究所奶牛营养学北京市重点实验室,北京100081)
NRC(2012)[1]和NRC(1994)[2]已给出详细的各个生长阶段的猪和家禽的氨基酸营养需要量,NRC(2000)[3]也给出了生长肉牛的氨基酸需要量预测模型,而关于奶牛生长阶段的氨基酸营养需要还缺乏系统的研究。NRC(2001)[4]指出后备牛的蛋白和氨基酸需要量的确定可划分为四个阶段:断奶前、过渡期(2~4月龄)、4月龄~配种期、配种后~产犊前。研究后备母牛的氨基酸需要量有助于提升牛场的经济效益和母牛的生产力。但目前氨基酸限制性顺序及需要量的研究多集中在犊牛和成年牛,有关于生长阶段后备牛氨基酸需要量的研究鲜见报道,饲粮氨基酸需要量及其理想氨基酸模式还处于空白。本文对牛生长阶段的氨基酸营养研究现状做一综述,旨在为后备牛生长阶段的氨基酸限制性顺序及需要量模型的研究提供参考。
1 牛生长阶段的限制性氨基酸
1.1 牛生长阶段的主要限制性氨基酸
小肠是生长牛吸收氨基酸的主要场所。小肠中的氨基酸主要来源于饲粮中的瘤胃非降解蛋白和瘤胃微生物蛋白[5],所以饲粮氨基酸组成并不是决定牛生长阶段的氨基酸需要量和限制性顺序的唯一因素。当微生物蛋白的量不能满足牛的生长需要时,某些氨基酸也会成为牛的限制性氨基酸。Titgemeyer等(1990)[6]研究表明,当真胃灌注蛋氨酸(Met)时,生长牛的氮(N)沉积增加17%。此外,只有当灌注Met时,N沉积才增加,所以Met可能是生长公牛的第一限制性氨基酸。但是仅仅灌注Met的效果不及混合灌注氨基酸的效果显著。Greenwood等(1999)[7]和Löest等(2001)[8]研究表明,Met、组氨酸(His)、赖氨酸(Lys)、亮氨酸(Leu)和缬氨酸(Val)是生长荷斯坦公牛的限制性氨基酸。这些结果显示,生长牛的限制性氨基酸不止一种,相比于只有一两种关键氨基酸,到达真胃的氨基酸越丰富,牛生长性能越好。
1.2 牛生长阶段的氨基酸限制性顺序
目前对犊牛及生长公牛氨基酸限制性顺序的研究很多。表1汇总了近年来国内外关于牛生长阶段氨基酸限制性顺序的研究结果。结果表明,生长牛的氨基酸限制性顺序受品种、日龄、体重等多个因素影响。Lys和Met为牛生长阶段的前两种限制性氨基酸,苏氨酸(Thr)、色氨酸(Trp)和His可能是牛生长阶段的第三限制性氨基酸。
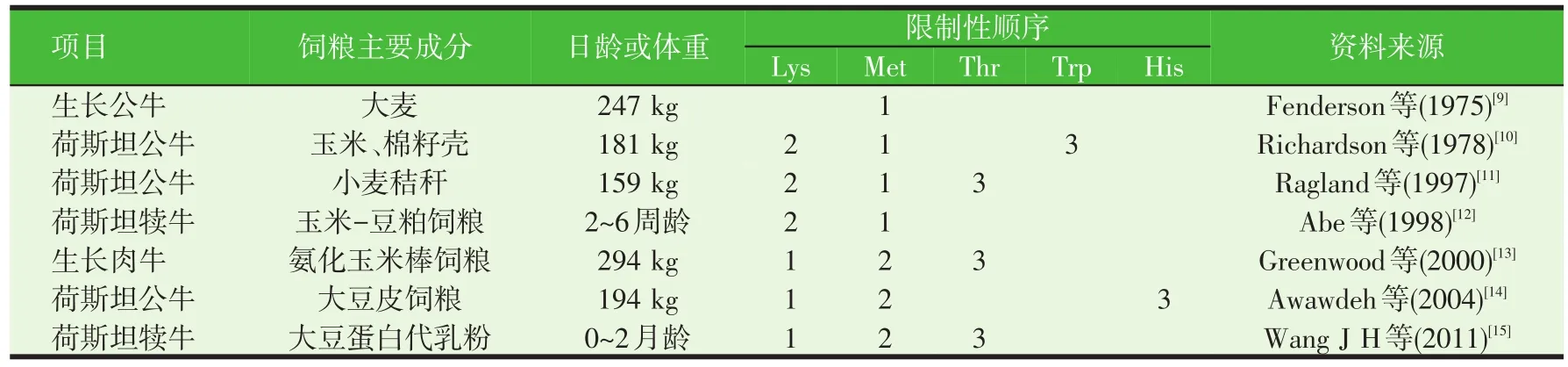
表1 牛生长阶段的氨基酸限制性顺序
大量研究表明饲粮的过瘤胃蛋白的含量是影响反刍动物生长阶段氨基酸限制性顺序的一个重要因素。大麦易被瘤胃降解,过瘤胃低,Fenderson等(1975)[9]研究表明,当生长公牛的饲粮含80%的大麦时,Met是第一限制性氨基酸。玉米蛋白的过瘤胃率较高,以玉米为基础饲粮的生长公牛灌注Lys,发现Lys可能是第一限制性氨基酸。当饲粮蛋白组成多为瘤胃降解蛋白时,到达小肠的氨基酸多为微生物蛋白,此时限制性氨基酸多与微生物蛋白的组成有关。Nimrich等(1970)[16]等研究表明以尿素为唯一氮源的生长绵羊,氨基酸限制性顺序为Lys、Met、Thr。
2 牛生长阶段的氨基酸平衡模式
2.1 氨基酸平衡
氨基酸平衡是保证氨基酸利用效率最大化的关键。实际生产中都会严格把控氨基酸的添加量,考虑多种限制性氨基酸之间的平衡关系,确定适宜的氨基酸添加比例,做到精准饲喂,以确保氨基酸平衡。饲粮中某些氨基酸的不足和过量都会使氨基酸之间失去平衡,从而影响畜禽的生长发育。Abe等(1999)[17]研究发现,向体重约为103 kg生长牛真胃内投喂0.333 g/kg BW的Met,育成牛会出现明显的中毒症状;Abe等(2001)[18]研究发现,150 kg的育成牛每天摄入64 g Lys时腹泻率明显增加。此外饲粮氨基酸平衡是降低饲养成本,节约蛋白饲料资源的一个首要手段。氨基酸平衡能保证最大程度发挥各个氨基酸的利用效率,从而降低饲粮蛋白水平,节约成本。
2.2 牛生长阶段的理想氨基酸模式(IAAP)的研究进展
近年来,理想氨基酸模式在成年奶牛的研究较多,对生长阶段反刍动物的氨基酸平衡模式的研究较少。Hill等(2008)[19]研究表明,5周龄前犊牛代乳料中Lys、Met和 Thr的适宜比例为 100∶31∶60;刁其玉[20]提出羔羊的氨基酸平衡模型为Lys、Thr和Trp比值为100∶50.5∶14.3。云强等(2011)[21]研究表明,8~16周龄犊牛的玉米-豆粕型开食料中适宜的Lys、Met比例为3.1∶1。
氨基酸平衡模式的构建除与饲粮和动物本身有关外,还受研究方法的影响。动物理想氨基酸模式的研究方法主要有析因法、计量效应法、屠体法和氨基酸部分扣除法。许多研究指出析因法和剂量效应法易受其他因素的影响,结果都存在一定的问题。陈正玲(2001)[22]研究用三种方法确定的肉鸭IAAP之间存在一定差异,屠体法测定的模式只能作为参考,剂量效应法测定的维持生长模式与AA部分扣除法确定的模式相对较为接近,AA部分扣除法确定的模式可能与肉鸭实际的IAAP更为接近。AA部分扣除法是基于析因法和剂量效应法两者优点于一体的准确测定IAAP的有效方法。通过扣除饲粮部分氨基酸观察动物的生长性能、饲料转化率、N沉积等指标,从而确定动物的氨基酸限制性顺序。Wang等(2010)[23]采用AA部分扣除法确定了猪的理想氨基酸模型,并作为NRC(2012)[1]修订猪IAAP的一个重要依据。但氨基酸扣除法在反刍动物IAAP的研究中应用不多,有两篇研究采用AA部分扣除法确定了犊牛和羔羊的IAAP。Wang J H等[15]采用氨基酸扣除法指出获得最大ADG时0~2和4~6周龄犊牛的Lys、Met和Thr需求比例为100∶35∶63和100∶27∶67。李雪玲等[24]采用氨基酸扣除法确定了60~90日龄和90~120日龄羔羊获得最大ADG、F/G和DP时,Lys、Met、Thr和Trp的适宜比例为100∶44∶42∶8和100∶41∶38∶11,并且采用屠体法验证发现二种方法确立的羔羊氨基酸适宜比例相近。
目前氨基酸部分扣除法已应用于单胃动物的氨基酸限制性顺序及平衡模式的研究,但此方法在反刍动物尤其在生长牛IAAP的研究方面的应用较少。由于反刍动物的特殊消化生理,该方法是否适用于不同阶段生长牛的IAAP研究需进一步验证,生长牛适宜的理想氨基酸模式研究方法也需进一步探索。
3 牛生长阶段氨基酸需要量的研究
由于反刍动物的特殊消化过程,评定反刍动物的氨基酸需要量不能根据其摄入的氨基酸量进行评定,要根据其进入小肠并被吸收的氨基酸量,也就是代谢氨基酸量来确定。表2和表3汇总了近年来国外学者对生长阶段牛的代谢氨基酸需要量的研究结果。我们可以看出品种、体重、日增重均是影响生长阶段牛的氨基酸需要量的因素。

表2 牛生长阶段代谢氨基酸的需要量

表3 生长肉牛Met的需要量
R.A.Zinn(1988)[33]根据表3中的研究成果拟合出了生长肉牛Met的需要量模型METR=1.956+0.029 2PG+0.029 0W(PG:蛋白增重,W:体重,R2=0.92),用此模型计算得的氨基酸需要量与其动物试验测定的生长肉牛氨基酸需要量相近。R.A.Zinn等(1998)[25]等将上述公式具体化,得出了依据体重和日增重确定Met需要量的模型:METR=(1.956+0.029 2×ADG[268-(29.4×0.055 7BW0.75×ADG1.097)÷ADG]+0.112×BW0.75),经过100头生长公牛的饲养试验验证得出本公式计算的小肠Met的提供量与实际测定量之间的相关性达0.99,此模型可用于计算肉牛生长阶段Met需要量。Montaño等(2016)[34]用192头公牛验证了NRC(2000)[3]氨基酸需要量计算模型的准确性,认为NRC(2000)[3]提供了可靠的计算饲粮提供给小肠代谢氨基酸量及生长牛代谢氨基酸的需要量的模型。但目前很少有研究证实此计算模型在生长牛增重关键时期的实用性。
由于瘤胃发酵的转化导致进入生长牛小肠的氨基酸的数量和种类不统一,计算到达小肠的氨基酸量非常困难,所以近些年有关建立不同阶段生长牛的氨基酸需要量模型研究很少。但模型的建立是确定各阶段反刍动物氨基酸需要量的基础,通过模型的拟合确立各阶段生长牛的氨基酸需要量是有待研究的重点。模型建立需要以大批量数据为基础,但目前生长阶段后备牛的氨基酸需要量还处于空白,所以研究生长后备牛的氨基酸需要量,为后续模型的建立提供基础刻不容缓。
4 牛生长阶段的微生物蛋白(MCP)的氨基酸组成及影响因素研究
瘤胃微生物以发酵产生的ATP和挥发性脂肪酸为能量和碳源,以氨基酸和氨为底物合成微生物蛋白质。微生物蛋白质是一种“高质量”的蛋白质,它会同过瘤胃蛋白质随食糜进人真胃和小肠,供动物体吸收和利用。大量研究表明,MCP可提供成年奶牛所需的绝大部分氨基酸;犊牛由于瘤胃发育不完善,MCP合成有限,微生物提供的氨基酸较少,其氨基酸主要由饲粮提供;生长牛的瘤胃发育基本完善,已具备基本的合成MCP的能力,所以MCP为生长牛氨基酸的另一主要来源。因此,MCP的氨基酸组成也可能会影响生长牛的氨基酸平衡及需要量。
大量研究证明,MCP的氨基酸组成与微生物的种类直接相关。瘤胃微生物主要分为细菌、原虫和微生物三种,此外还可根据其存在方式分为液相菌(游离于瘤胃液)和固相菌(附着于饲料颗粒)。表4汇总了近几年研究中的瘤胃微生物蛋白的主要氨基酸组成。从表中可以看出不同群系MCP主要氨基酸组成存在一定差异,Rodríguez等[35]和周勃等[36]结果发现固相菌中Lys含量高于液相菌;王洪荣等(2009)[37]研究发现,原虫、细菌和微生物的氨基酸组成存在一定的差异,原虫的Val含量高于细菌和微生物,细菌和微生物的Lys含量高于原虫。

表4 瘤胃MCP的氨基酸组成
有研究表明,微生物区系的变化是造成瘤胃微生物中某些氨基酸含量和比例差异较大的原因。而瘤胃微生物区系可能受饲粮等其他因素的影响。Vogel等(1983)[39]研究表明,饲粮对瘤胃微生物的氨基酸组成没有影响;而Erasmus等(1994)[40]、王洪荣(1998)[41]的研究表明瘤胃微生物的氨基酸组成会受饲粮的影响。迄今,有关MCP的氨基酸的研究已有很多,但结果不一,瘤胃微生物氨基酸组成是否会改变,改变的原因及机理仍不清楚。作为牛生长阶段氨基酸主要来源之一,MCP的氨基酸组成对其氨基酸平衡及需要量的影响未见研究。所以以MCP氨基酸组成研究为切入点,研究其对生长牛氨基酸平衡的影响是生长牛氨基酸营养研究的另一个创新点。
5 小结
奶牛理想氨基酸模型的建立已取得一定的进展,但大多局限于成年奶牛和犊牛,后备牛理想氨基酸模型的研究还处于空白。饲粮是决定后备牛氨基酸平衡的一个重要因素,研究饲粮对氨基酸需要量的影响及生长牛小肠氨基酸的吸收及转运机制对氨基酸平衡模式的构建有重要意义。此外,还需把MCP的氨基酸组成作为重点,加强对MCP氨基酸组成及改变机理的研究。最终以饲粮和MCP氨基酸含量和组成为基础,筛选合适的理想氨基酸模型的构建方法,成功构建后备牛理想氨基酸模式。目前的氨基酸需要量研究大多以动物血液或肉中的氨基酸量为基础,反向倒推氨基酸需要量;或者以饲粮和小肠食糜氨基酸含量为基础确定氨基酸需要量。MCP的氨基酸组成很少被考虑,所以MCP是否对生长牛氨基酸平衡和需要量有影响也是建立生长牛理想氨基酸模型需考虑的另一主要因素。
