奶牛精子变形期间Pld6基因表达及蛋白序列分析
2019-01-02李小俊陈晓丽赵毅强路永强
李小俊,李 欣,陈晓丽,赵毅强,路永强,王 栋*
(1.中国农业科学院 北京畜牧兽医研究所,北京 100193; 2.中国农业大学 生物学院,北京100193;3.北京市畜牧总站,北京 100107)
线粒体是细胞内通过氧化磷酸化产生能量的重要细胞器[1-4],可根据内外刺激发生融合、裂变、运输和自噬等复杂变化[5-7]。哺乳动物精子变形过程中,线粒体在数量、形状、结构等方面发生了显著变化,线粒体嵴贴附在线粒体外膜内侧,增大了线粒体内弥散的空泡化基质空腔,并最终形成月牙状的线粒体,螺旋式包裹在精子鞭毛中段,形成线粒体鞘,为精子提供生存、运动所需的ATP[8-15],对精子尾部骨架形态和功能发挥重要作用。然而,在精子形成过程中线粒体鞘形成的调节过程仍有待确定。磷脂酶D家族成员6(Pld6/MitoPld)是物种间保守的磷脂酶D蛋白超级家族成员,定位于线粒体外膜,可以水解心磷脂(cardiolipin, CL),产生信号分子磷脂酸(phosphatidic acid, PA),诱导线粒体融合[7,16-19],Pld6缺陷会导致小鼠线粒体错误定位于中心体周围,影响精子形成正常形态[20]。甘油激酶样蛋白1(glycerol kinase-like 1, Gykl1)和甘油激酶2(glycerol kinase 2, Gyk2)可以与Pld6相互作用,激活线粒体聚集所依赖的Pld6和PA,Gykl1和Gyk2缺失会使Pld6和PA激活受阻,导致精子线粒体形态异常、线粒体鞘无序排列和精子尾部缺陷,并使精子ATP的产生失控[21]。由此可见,Pld6对于小鼠形成正常功能的精子线粒体鞘至关重要,而该基因在精子形成过程中的表达尚不清楚,对于牛的研究未见报道。精子线粒体形态和功能的正常形成,直接影响到种公牛的精液品质和精子活力,进而影响其繁殖力。本研究在奶牛圆形、长形精子细胞和附睾尾精子转录组研究的基础上,检测了精子变形过程中3种不同形态细胞间差异表达基因Pld6的表达水平,并预测了牛Pld6蛋白结构,为进一步研究该基因在牛精子形成中的重要作用提供理论依据。
1 材料与方法
1.1 试剂
HistoGene LCM Frozen Section Staining Kit(Life technology);SG高纯总RNA提取试剂盒、Thermo First cDNA Synthesis Kit、2×SG Green qPCR Mix (SinoGene)。
1.2 牛睾丸及附睾组织的采集
选取3头健康状况良好、15~17月龄、体况相近、有正常生育能力的荷斯坦公牛,用灭菌手术剪采集睾丸和附睾组织,分别置于保鲜袋中,标注后迅速置于冰盒中运回实验室。
1.3 试验方法
1.3.1 牛附睾尾精子的采集及活力检测 用手术剪从睾丸上剥离出附睾尾组织,并置于装有PBS的RNase-free离心管(50 mL)中,标记后剪碎,于4 ℃静置30 min,使精子从附睾尾中充分游离释放,小心吸取上清,收集附睾尾精子。取1 μL上述精液,稀释10倍后使用血细胞计数板,在400×物镜下统计直线运动的精子占总精子数量的百分率,以3个视野的平均数作为精子活力等级,活力高于50%的精子用于后续试验。
1.3.2 牛圆形及长形精子细胞的获取 将新鲜离体睾丸剪成1.0 cm×0.6 cm×0.4 cm的组织块,用OCT包埋后置于液氮中速冻2~3 min,迅速置于预冷的-20 ℃冷冻切片机内平衡30 min,将样品固定后,修片厚度为20 μm,正式切片时厚度调为5 μm,用PEN 2.0膜玻片(RNase-free)贴片后,置于-80 ℃储存备用或直接使用试剂盒染色。然后采用Leica LMD7000系统确认染色切片中的圆形、长形精子细胞,沿目标细胞边缘划线并切割。切割细胞因自身重力落到含细胞裂解液的收集管盖子中,切割完毕后取下收集管,加入75 μL裂解液,按紧盖子,反复颠倒2 min,收集细胞,-80 ℃或液氮保存备用。每张膜玻片的切割时间控制在40 min以内[22]。
1.3.3 样本总RNA提取及cDNA合成 参照TRizol使用说明书提取细胞样品总RNA,用超微量分光光度计检测总RNA浓度和纯度,按照反转录试剂盒操作说明,以Oligo(dT)18为引物,将质量合格的RNA反转录合成cDNA。
1.3.4 差异基因筛选 通过RNA-seq对奶牛圆形、长形精子细胞及附睾尾精子进行转录组测序,采用DESeq进行基因差异表达分析,比较圆形、长形精子细胞及附睾尾精子的转录组数据,并选取|log2Ratio|≥1和q<0.05的基因作为差异表达基因,利用Blast2GO,得到每个基因对应的GO条目,通过差异基因GO富集分析,得到这些差异基因显著相关的生物学功能,并挑选与线粒体相关的条目,然后,根据基因功能筛选差异基因用于qPCR分析。
1.3.5 引物设计与合成 根据NCBI数据库牛Pld6基因的mRNA序列(登录号:NM_001271990.1)和beta-肌动蛋白基因(β-actin)序列(登录号:NM_173979.3),使用NCBI在线工具设计PCR引物。引物信息如表1所示。
表1qPCR引物信息
Table1Theinformationofprimersdesigned
1.3.6 差异基因的实时荧光定量PCR(qPCR)分析 以合成的cDNA为模板,按照2×SG Green qPCR Mix反应体系,以β-actin基因为内参进行qPCR扩增。设置3个生物学重复,使用Excel统计附睾尾精子及活力;用Prizm4软件通过比较Ct方法对基因进行表达量分析。使用“平均值±标准误”表示,P<0.05表示差异显著;P<0.01差异极显著。
1.3.7 Pld6生物信息学分析 在Uniprot下载牛Pld6蛋白的氨基酸序列,通过ProtParam软件(https://web.expasy.org/protparam)分析该蛋白质分子组成、相对分子质量、理论等电点及稳定性等参数,使用TMHMM软件(http://www.cbs.dtu.dk/services/TMHMM-2.0/)进行跨膜区域分析,通过SOPMA软件(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html) 和Swiss_model软件(https://www.swissmodel.expasy.org/) 预测蛋白二级和三级结构,通过NCBI进行BLAST搜索并下载已知同源序列,使用DNAMAN进行多序列比对分析并构建系统进化树。
2 结 果
2.1 奶牛精子细胞采集
通过浮游法获取附睾尾精子,其精子形态如图1A所示,其头部呈椭圆形,轮廓规则,几乎看不到畸形精子。活力统计结果:1、2、3号样品的精子数分别为1.28×109、1.58×109、1.24×109,其中,直线运动精子占精子总数的百分率分别为54%、55%、61%,可以用于后续研究。
奶牛睾丸组织冰冻切片染色后,显微镜下可见圆形精子细胞(图1B),细胞核偏向细胞一侧或居中;长形精子细胞呈长条状、两头钝圆(图1D)。LCM切割前圆形精子细胞见图1B(箭头指出并红色圈出的细胞);LCM切割后的圆形精子细胞见图1C;LCM切割前长形精子细胞见图1D(箭头指向并绿色圈出的细胞);LCM切割后的长形精子细胞见图1E,细胞样品收集情况见图1F和表2。
2.2 奶牛圆形、长形精子细胞及附睾尾精子总RNA的制备及含量分析
表3显示,所有细胞样本总RNA的OD260 nm/OD280 nm为1.91~1.98,纯度较高。圆形精子细胞总RNA量为2 000~2 650 ng,长形精子细胞总RNA量为2 700~3 200 ng,而附睾尾精子总RNA含量为2 300~3 700 ng。

A. 普通光学显微镜下附睾尾精子形态(200×);B.切割前圆形精子细胞(箭头指出并红色圈出的细胞)(400×);C.切割后圆形精子细胞(与图B对应圈出的细胞)(400×);D.切割前长形精子细胞(箭头指出并绿色圈出的细胞)(400×);E.切割后长形精子细胞(与图D对应圈出的细胞)(400×);F.激光显微切割仪切割后收集到的细胞(150×)A. Sperm morphology of epididymis tail under general optical microscope(200×); B.Photo before cutting round spermatids (the arrows point and circle the cells in red) under the microscope of LCM(400×);C.Photo after cutting round spermatids under the microscope of LCM(the corresponding cells to figure B circled)(400×);D.Photo befor cutting elongated spermatids (the arrows point and circle the cells in green) under the microscope of LCM(400×);E.Photo after cutting elongated spermatids (the corresponding cells to figure D circled) under the microscope of LCM(400×);F.The collected spermatids under the microscope of LCM(150×)图1 精子细胞的获取Fig.1 Collection of spermatogenic cells
表2收集到的圆形及长形精子细胞数
Table2Thenumberofroundandelongatedspermatids

样本Sample圆形精子细胞数(R)Round spermatids No.长形精子细胞数(E)Elongated spermatids No.16 1886 02926 0596 122310 25710 551
表3总RNA浓度检测
Table3TheconcentrationdetectionofRNA
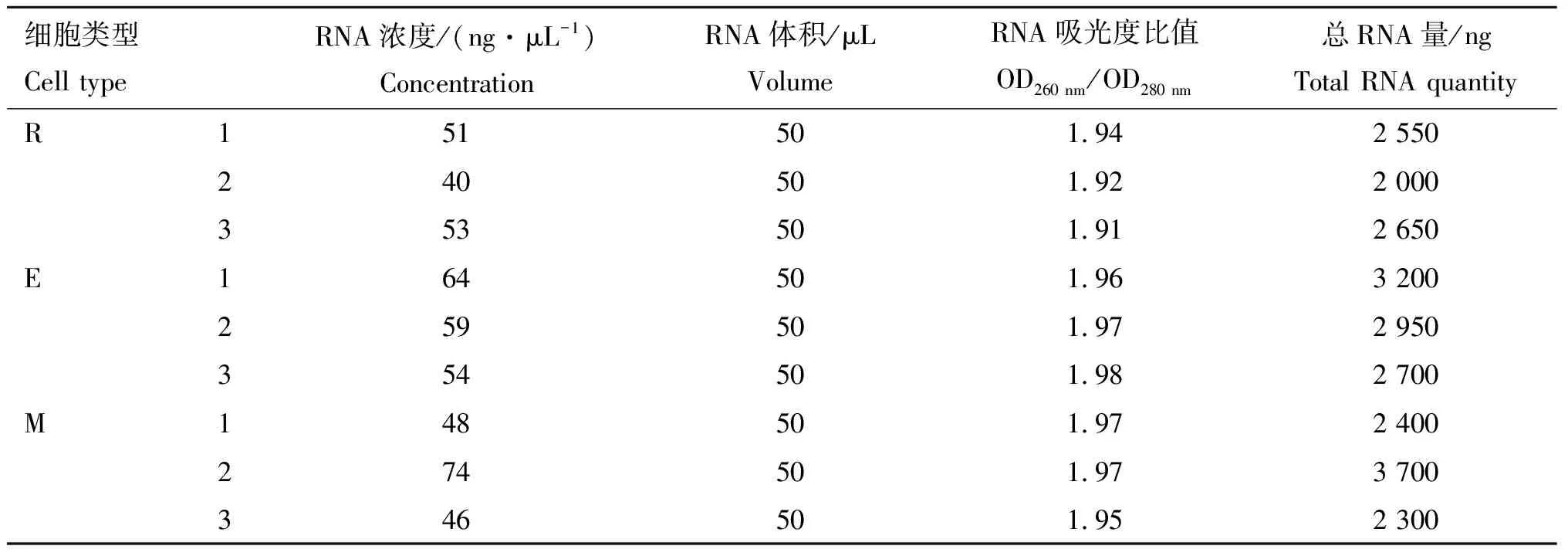
细胞类型Cell typeRNA浓度/(ng·μL-1)ConcentrationRNA体积/μLVolumeRNA吸光度比值OD260 nm/OD280 nm总RNA量/ngTotal RNA quantityR151501.942 550240501.922 000353501.912 650E164501.963 200259501.972 950354501.982 700M148501.972 400274501.973 700346501.952 300
R.圆形精子细胞;E.长形精子细胞;M.附睾尾精子。1、2、3分别为3类细胞的生物学重复
R. Round spermatid; E. Elongated spermatid;M. Epididymal sperm.1,2,3. The 3 biological duplication of 3 cell types
2.3 差异基因筛选
通过DESeq进行基因差异表达分析,共发现5 349个基因在精子形成阶段差异表达(|log2Ratio|≥1,q<0.05),其中,圆形到长形精子的差异表达基因为994个(246个上调,748个下调);长形到附睾尾精子差异表达基因为4 771个(1 613个上调,3 158个下调),根据基因注释,将这些基因进行富集分析,发现了与线粒体及精子变形相关的功能条目及差异基因(表4),其中,Pld6在圆形、长形和附睾尾精子中的表达量(FPKM值)分别为492.00、1 870.67和0.00,即在圆形变形为长形精子细胞的过程中表达量升高,为圆形精子细胞的3.8倍,而在附睾尾精子中表达量降低为0,且该基因富集于线粒体融合(mitochondrial fusion)功能条目中,根据其测序数据及注释的重要功能,筛选该基因进行后续分析研究。
表4与线粒体相关的功能条目及富集的基因数
Table4Mitochondrial-relatedGOtermsandenrichedgenenumber

GO条目GO term功能描述Description差异基因数Gene numberGO:0007005线粒体组织Mitochondrion organization99GO:0008053线粒体融合Mitochondrial fusion8GO:0000422线粒体降解Mitochondrion degradation24GO:0007007线粒体内膜Inner mitochondrial membrane organization4GO:0051646线粒体定位Mitochondrion localization5GO:0007006线粒体膜Mitochondrial membrane organization14GO:0042775线粒体ATP合成耦合电子传递Mitochondrial ATP synthesis coupled electron transport16GO:0000266线粒体分裂Mitochondrial fission13
2.4 Pld6在奶牛圆形、长形精子细胞和附睾尾精子中的转录表达检测
Pld6基因的qPCR检测结果(图2)显示,Pld6基因在圆形到长形精子细胞期间表达量升高,而在后期的精子成熟过程中表达量降低,与测序数据一致。
2.5 Pld6蛋白的特征分析
ProtParam分析表明,Pld6蛋白由220个氨基酸组成,分子式为C1128H1799N331O306S8,相对分子质量为25 150.21,理论等电点为9.77,不稳定指数为61.43,属于不稳定蛋白。通过SOPMA对其二级结构分析显示,该蛋白含有39.55%α螺旋,29.09%随机卷曲,20.91%延伸链,10.45%β折叠(图3),TMHMM对其跨膜结构预测发现,第1~9氨基酸位于膜外,10~32氨基酸是跨膜区域,33~220氨基酸位于膜内,其中跨膜区域为该蛋白在线粒体外膜定位区域(图略)。对其三级结构进行预测(图4),α螺旋区域位于外围,β折叠区域被α螺旋区域包围在内侧,呈喇叭状。

R.圆形精子细胞;E.长形精子细胞;M.附睾尾精子。标有不同大写字母和不同小写字母者分别表示组间差异极显著和差异显著(P<0.01,P<0.05)R.Round spermatid; E.Elongated spermatid; M.Epididymal sperm.Those marked with different capital letters and different lowercase letters respectively indicate extremely significant differences (P<0.01) and significant differences (P<0.05) among different groups图2 精子变形期间Pld6基因qPCR检测结果Fig.2 Relative expression of Pld6 gene during spermiogenesis
2.6 奶牛Pld6蛋白同源性及进化分析
利用NCBI的BLASTp对牛Pld6进行同源搜索,结果表明,牛(Bostaurus)与小鼠(Musmusculus)、黑猩猩(Pantroglodytes)、人(Homosapiens)、光滑爪蟾(Xenopuslaevis)、非洲爪蟾(Xenopustropicalis)和斑马鱼(Daniorerio)的同源性分别为85%、85%、84%、60%、60%和55%。用DNAMAN对下载蛋白序列的多重比对(图5)发现,这些蛋白有一个明显的Pld家族区域H(X)K(X4)D (HKD),是该蛋白的活性区域。对这7个蛋白建立系统发育树(图6),Pld6进化分为鱼类、两栖类和哺乳类3个分支,符合动物分类学地位。
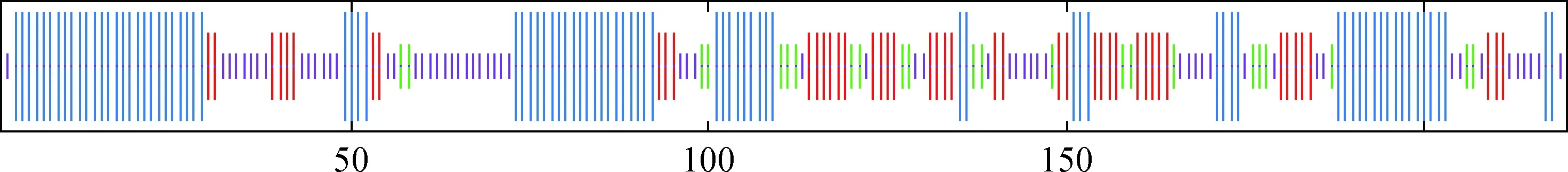
蓝色.α螺旋;绿色.β折叠;红色.延伸链;紫色.随机卷曲Blue.Alpha helix; Green. Beta sheet; Red. Extended strand; Purple.Random coil图3 Pld6蛋白的二级结构预测结果Fig.3 Secondary structure prediction of Pld6 protein

A.α螺旋;B.β折叠;C.延伸链;D.随机卷曲A.Alpha helix; B. Beta sheet; C. Extended strand; D. Random coil图4 Pld6蛋白的三级结构预测结果Fig.4 Prediction of tertiary structure of Pld6 protein
3 讨 论
结构是正常功能充分发挥的基础,精子变形期间尾部正常结构的形成,对精子活力及受精能力形成具有重要作用[13-15,22-24],本研究通过对精子形成过程中大量基因的差异表达分析,发现了与精子尾部中段线粒体分布和形态结构相关的Pld6基因,又利用后续qPCR结果验证了其差异表达的真实性,并通过对该蛋白可能的结构和功能分析,对该基因与精子尾部结构和功能的关系进行了科学推断。
哺乳动物精子形成过程中,精子细胞形态发生了复杂变化[8-12],为探究精子变形的机制,本实验室前期对奶牛圆形、长形精子细胞及附睾尾精子进行了转录组测序,测序数据分析结果表明,Pld6基因在圆形到长形精子细胞发育过程中表达量升高,在附睾尾精子中未检测到其转录表达,通过GO分析,将该基因富集在mitochondrial fusion条目。研究表明,线粒体形态通过线粒体融合和裂变的动态平衡维持,Pld6基因参与该平衡的维持[25-26],可能影响精子尾部特异的线粒体鞘形态的形成和维持。而Pld6激活失败或缺陷会使小鼠线粒体错误定位于中心体周围,导致精子形成停滞,或使精子线粒体形态异常、线粒体鞘无序排列和精子尾部缺陷[20-21],因此,推测该基因还对牛精子线粒体鞘形成发挥重要作用。而目前尚未见到对牛Pld6功能的研究,为进一步探索该基因功能,本研究对牛Pld6蛋白质序列进行了生物信息学分析,发现该蛋白在不同物种间具有保守的H(X)K(X4)D (HKD)结构域,并呈现酶活性区域特征,推测在圆形精子向长形精子发育过程中,Pld6基因表达产物可通过该功能域催化CL产生PA,并进一步通过该信号分子,诱导精子细胞线粒体向尾部中段聚集和相互融合,促进了精子尾部特异形状和功能的形成,该推断可通过相关文献[7,27-29]得到验证。研究表明,小鼠Pld6定位于线粒体外膜[20,30],其三级结构呈喇叭状,该蛋白活性区域为β折叠,处于喇叭状结构内部[31],这与本研究预测的牛Pld6蛋白结构相似。同时,本研究关于该基因蛋白序列同源物种的聚类分析表明,该蛋白通过长期进化,已分化为鱼类、两栖类和哺乳类3个进化方向,其中,牛与小鼠关于Pld6蛋白具有最接近的进化趋势。聚类分析结果为将来采用比较组学技术,充分利用小鼠研究成果,指导牛上深入开展该基因及蛋白在精子形成中对线粒体鞘形成的作用提供一定的理论基础,对提高种公牛繁殖力有积极意义。
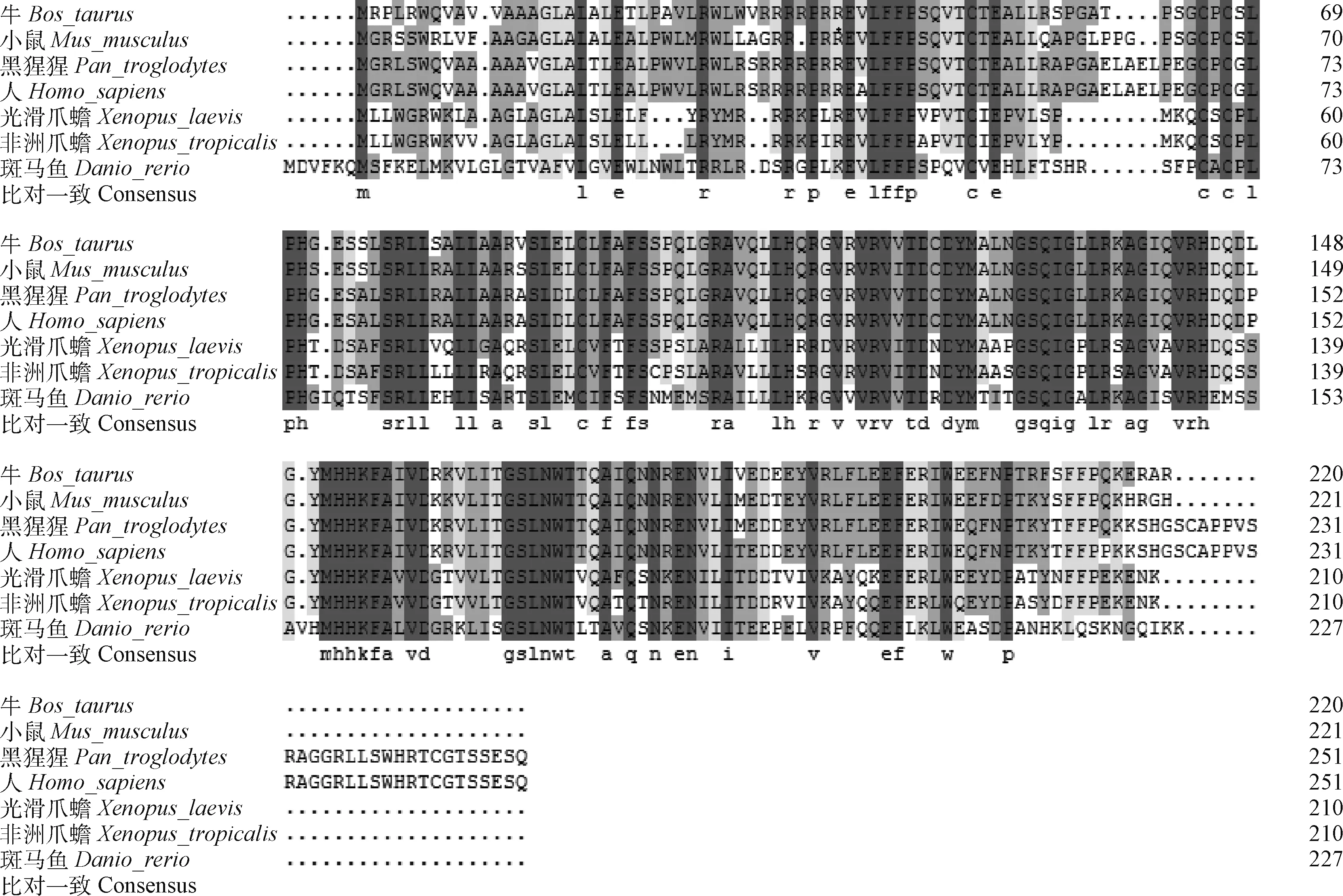
图5 不同物种Pld6蛋白序列比对结果Fig.5 The sequence alignment of Pld6 protein among different species

图6 Pld6蛋白系统发育树分析Fig.6 Phylogenetic tree analysis of Pld6 protein
4 结 论
本研究根据小鼠研究推断牛精子变形过程中,Pld6基因表达增多可诱导线粒体向精子细胞尾部中段聚集,促进了线粒体鞘的正常形成,对确保牛精子活力及受精能力具有重要作用,为进一步研究Pld6基因功能,提供了重要基础。关于Pld6基因的表达调控及其产物对精子中线粒体形态形成和功能发挥的作用,还需要进一步的研究验证。
