亚高山森林自然与人工恢复对土壤涵水能力的影响
2018-12-20祁凯斌黄俊胜杨婷惠包维楷庞学勇
祁凯斌,黄俊胜,杨婷惠,包维楷,庞学勇,*
1 中国科学院山地生态恢复与生物资源利用重点实验室,生态恢复与生物多样性保育四川省重点实验室,中国科学院成都生物所,成都 610041 2 中国科学院大学,北京 100049
森林是陆地生态系统的主体,通过林冠层、林下植被层、枯枝落叶层以及土壤的复杂结构,在水土保持和水源涵养等功能中发挥巨大的作用,其中地上部分的持水量仅占森林生态系统水源涵养能力的15%,土壤层是森林生态系统水源涵养的主体[1]。森林土壤具有大量的土壤孔隙,降雨能够沿着孔隙下渗,成为土壤贮水和地下径流,进而减少地表径流,从而起到涵养水源和保持水土的作用[2]。对退化生态系统进行人工恢复是当前一种较为推崇的林业实践措施,特别是在采伐迹地上再造林被喻为能迅速恢复生态系统生产力[3]。但越来越多的研究表明,单一树种造林,林地结构单一,生态位高度重合,导致人工林林下物种单一、土壤肥力退化等[4- 6]。与自然恢复途径相比,人工恢复可引起林地特征(物种、林木密度、根系特征等)差异,如树种本身差异导致凋落物分解的特性的不同,进而影响有机物归还速率,从而影响土壤中有机质的含量与质量[7],而有机质含量的多少决定了土壤是物理结构(如孔隙度等),土壤物理结构是土壤持水能力的基础[8]。另一方面,以单一树种造林形成的林地结构单一,缺乏林下植被,地表裸露导致表层土壤受径流冲刷而流失[9]。再者,自然与人工恢复途径也可能导致土层中根系的分布特征发生明显地变化,根系穿插决定着土壤中孔隙的分布与大小孔隙的分配,进而影响土壤的持水能力[10]。因此,随着恢复的进程,由于恢复途径不同以及林龄的差异,各林型树种生物学特性与林分结构存在不同,进而可能导致不同恢复途径下土壤水源涵养功能的差异。
川西亚高山针叶林是西南林区的主体,集中分布在岷江、金沙江、雅砻江等流域, 是长江上游水源涵养的重要组成部分[11]。随着20世纪40年代川西亚高山原始森林的大规模采伐,随后在采伐的斑块上进行了以云杉(Piceaspp.)为主要树种的人工造林[12];同时,桦木(Betulaspp.)等阔叶先锋树种的自然更新也在大规模进行[13],直至1998年,天然林资源保护工程正式启动,川西森林全面禁伐封育。在经历大规模采伐、人工更新及封育后,川西亚高山森林景观与格局都发生了深刻变化,不同恢复阶段的人工林、自然恢复次生林分布其中,为我们研究不同恢复途径下土壤生态功能变化提供了天然的试验场地。截至目前,关于该区域森林植被水源涵养功能的研究内容主要涉及林冠层截留[14]、地被物持水特性[15]、森林蒸发散[16]、土壤入渗[17]等,但对于自然与人工恢复途径的差异而影响土壤水源涵养功能的研究较少有报道[18]。该区域是国家生态功能中水源涵养和生物多样性规划的主体功能区,但目前对亚高山区不同恢复途径下土壤水源涵养功能还十分不清楚,进而深远影响着区域人工林生态系统恢复实践效果与功能状况的评估。
本文选择川西米亚罗林区系列林龄的人工恢复云杉林和自然恢复以桦木为主的针阔混交林,分析两种恢复途径林地的土壤水源涵养功能,揭示不同恢复途径林型特征与土壤水源涵养功能之间的关系,提出适宜的恢复措施和改善林地水环境,为加快川西退化生态系统功能提升提供重要的科学依据。
1 研究地区与研究方法
1.1 研究区域概况
研究区位于四川省阿坝州理县米亚罗林区(31°24′—31°55′N,102°35′—103°4′E,海拔2200—5500 m),地处青藏高原东缘的褶皱带最外缘部分,海拔高差悬殊,形成明显的高山峡谷地貌。该区属于高山气候,年平均温度6—12℃,极端最高温度32℃,极端最低温度-16℃,年无霜期约200 d,年均降雨量700—1400 mm,年蒸发量1000—1900 mm,土壤类型为山地暗棕壤。研究地段选择在米亚罗林区经过大规模砍伐、人工更新及实施封育后,不同恢复阶段的人工云杉林(Piceaasperata)、次生红桦林(Betulaalbo-sinensis)聚集分布地区。林下主要灌木有箭竹(Fargesiaspathacea)、悬勾子(Rubuscorchorifolius)、疏花槭(Acerlaxiflorum)、红毛五加(Acanthopanaxgiraldii)、散生枸子(Cotoneasterdavaricatus)、扁刺蔷薇(Rosasweginzmaii)、紫花卫茅(Euonymusporphyreus)、川滇高山栎(Quercusaguifolioides)等。主要草本有唐松草(Thalictrumuncatum)、蟹甲草(Cacaliaroborowskii)、冷蕨(Cystopterisfragils)、苔草(Carexsp.)、柳叶菜(Epilobiurnlaetum)和禾本科多属等[6]。
1.2 研究方法
1.2.1 样地设置与样品采集
在前期研究的基础上[19],根据恢复途径、林龄、海拔等条件选取了米亚罗林区大规模砍伐后,不同恢复途径下系列恢复年限(20 a、30 a、40 a、70 a)人工云杉林(Piceaasperata),和次生红桦林(Betulaalbo-sinensis),2类典型森林植被作为研究对象,林地年龄根据当地森工局的采伐历史,结合现场用生长锥钻取树芯粗略确定,再由带回实验室的树芯进行精确确定。选择立地条件大致相似的地段设置样地,尽量最小化土壤空间异质性因子,各样地主要基本特征见表1。
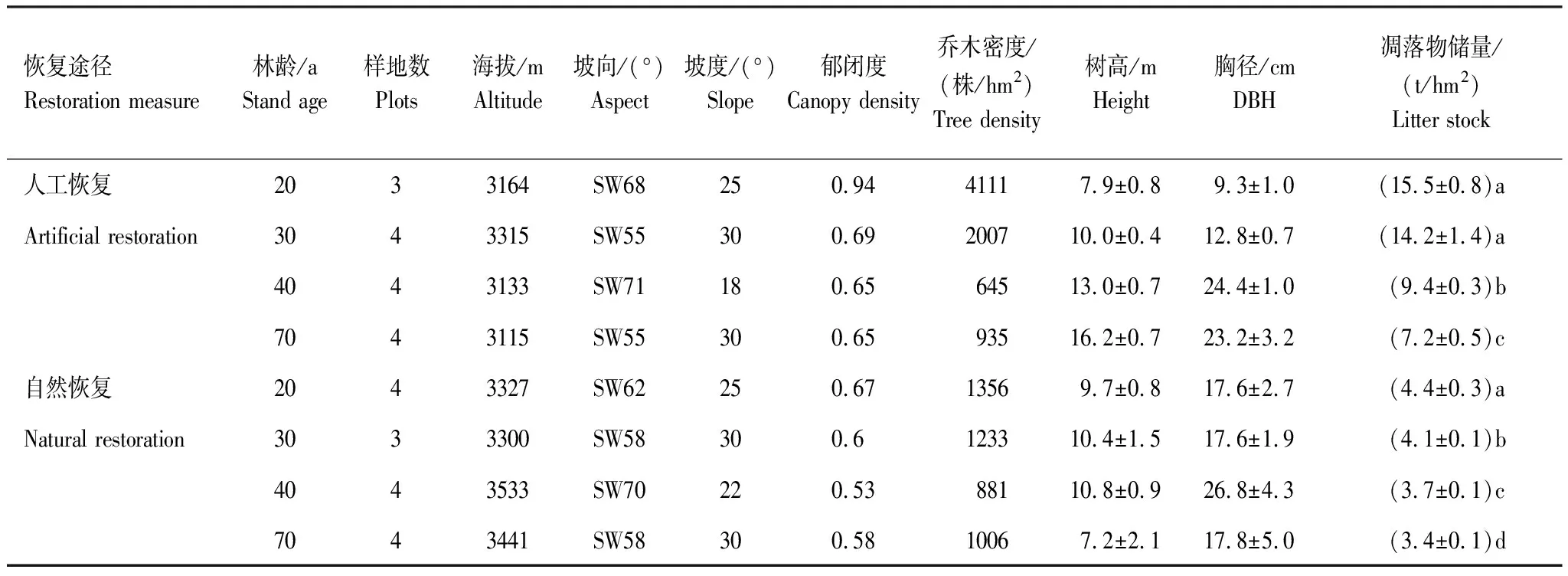
表1 研究地区不同恢复途径样地基本特征
小写字母不同表明其同一恢复途径不同年龄间差异达到显著水平P=0.05
2015年10月进行土壤样品的采集,于每个林地随机布设至少3个20 m×20 m标准样地,样地之间间隔至少50 m,在每个样地顺坡上、中、下各设置2个1 m×1 m小样方,先收集地表凋落物,然后在各点挖掘1个土壤剖面,用环刀法采集0—10、10—20 cm及20—40 cm 3个层次土壤样品,带回实验室内测定土壤孔隙和水分等物理性质,凋落物带回实验室65℃烘干至恒重并称重,计算凋落物储量;另在各采样点,用直径为5 cm的土钻分土层(0—10、10—20 cm和20—40 cm)采集土壤样品,同一样方的同一土层充分混合均匀后,每份取1000 g混合样,将野外采集的土壤样品中砾石及粗木质有机残体等捡除,并将小于2 mm的细根单独挑出,收集于信封内,带回实验室65℃烘干至恒重并称重,计算细根生物量。处理后的土样放入塑料袋中带回,置于阴凉通风处风干,用于测定土壤有机碳(TOC),土壤TOC含量采用元素分析仪(Vario Macro Analyzer,德国)测定。
1.2.2 土壤持水特性测定
土壤持水特性使用环刀法进行测定,将装有原状土壤的环刀在水中浸泡12 h称重(ms1,g),计最大持水量(Cmax,g/cm3),然后放置于干砂上2 h,此时环刀中土壤的非毛管水已全部流出,称重(ms2,g),计为毛管持水量(Ccap, g/cm3)。该方法详见《森林土壤定位研究方法》[20],持水量的计算公式如下:
(1)
(2)
Cnon=Cmax-Ccap
(3)
式中,ms0、ms1、ms2(g)分别为环刀内土壤干重、浸泡12 h后的饱和重量、失去非毛管水后的重量。v(cm3) 为环刀容积;Cmax、Ccap、Cnon(g/cm3)分别为土壤的最大持水量、毛管持水量和非毛管持水量。单位换算后,求出不同恢复途径下各林地相应土层的持水量(t/hm2)。
1.3 数据处理
对同一土层相同恢复途径林地土壤持水量(最大持水量、毛管持水量和非毛管持水量)和孔隙度等指标随年龄变化采用单因素方差分析(One-way ANOVA),同一恢复途径不同林龄的凋落物储量差异也采用单因素方差分析(One-way ANOVA),对于同一土层相同林龄不同恢复途径下土壤持水量和孔隙度等指标的差异性采用t检验,土壤持水特性指标与土壤理化性质、凋落物储量、根系生物量等指标的相关分析采用Person相关分析法。实验数据采用SPSS 20.0进行处理分析,制图使用Origin 9.1。
2 研究结果
2.1 林地及土壤基本性质
从表2可以看出,云杉人工林各土层平均容重变化范围在0.8—1.4 g/cm3,平均为1.02 g/cm3,容重随着林龄的增加而增加。次生林各土层平均容重变化范围在0.6—1.0 g/cm3,平均为0.79 g/cm3,容重随着林龄的增加而降低。在相同林龄条件下次生林各个土层容重低于人工云杉林。同时,随着土层的增加,人工云杉林和次生林的容重都在显著增加。
与此对照,土壤总孔隙度与土壤容重在两种恢复途径的各样地中呈相反趋势(表2),随着林龄的增加,次生林的同层次土壤总孔隙度要大于人工云杉林(表2)。人工云杉林土层总孔隙度在49.2%—70.9%之间变化,土壤总孔隙度随着林龄的增大而减小。次生林各土层总孔隙度在61.9%—78.9%之间变化,随着林龄的增加,土壤总孔隙度增加,同时随着土层的增加两种途径的土壤总孔隙度都在降低。人工云杉林和次生林土壤毛管孔隙度分别介于34.5%—44.3%和34.2—53.3%之间,随着林龄的增加,人工云杉林和次生林土壤毛管孔隙度均呈增加趋势,但次生林增加的幅度略大于人工云杉林。而土壤非毛管孔隙在人工云杉林和次生林各土壤层间差异不明显,人工恢复各层的土壤非毛管孔隙度介于16.6%—36.2%之间,随着林龄的增加,0—10 cm土层非毛管孔隙度降低,其余土层变化不明显。自然恢复各阶段的土壤非毛管孔隙度介于4.2%—30.9%之间,各土层土壤非毛管孔隙度随林龄的变化趋势不明显。
自然含水率表征林地瞬时水分状态,由表2可以看出,自然与人工恢复途径下,土壤的自然含水率差异明显,随着林龄的增加自然恢复的土壤自然含水率要大于人工恢复林。人工恢复的各土层自然含水率变化范围在27.8%—47.1%,随着林龄的增加,自然含水率在波动性降低。自然恢复的各土层自然含水率变化范围为31.0%—64.5%,随着林龄的增加,自然含水率在波动性增加。
人工恢复云杉林的土壤有机碳变化范围为14.7—85.5 g/kg,且随着年龄的增加而减少,而自然恢复的有机碳变化范围为24.0—93.1 g/kg,随林龄的变化趋势不明显,随着林龄的增加,自然恢复的土壤有机碳含量要大于人工恢复(表2)。细根生物量在人工云杉林和次生林之间存在差异,随着林龄的增加,次生林的细根生物量大于人工恢复云杉林。人工恢复云杉林细根生物量在0.54—1.98 t/hm2之间变化,随着林龄的增加细根生物量在降低。自然恢复次生林细根生物量在0.58—2.33 t/hm2之间变化,随着林龄的增加细根生物量也在增加。同时随着土层的增加,两种途径的细根生物量都在降低。
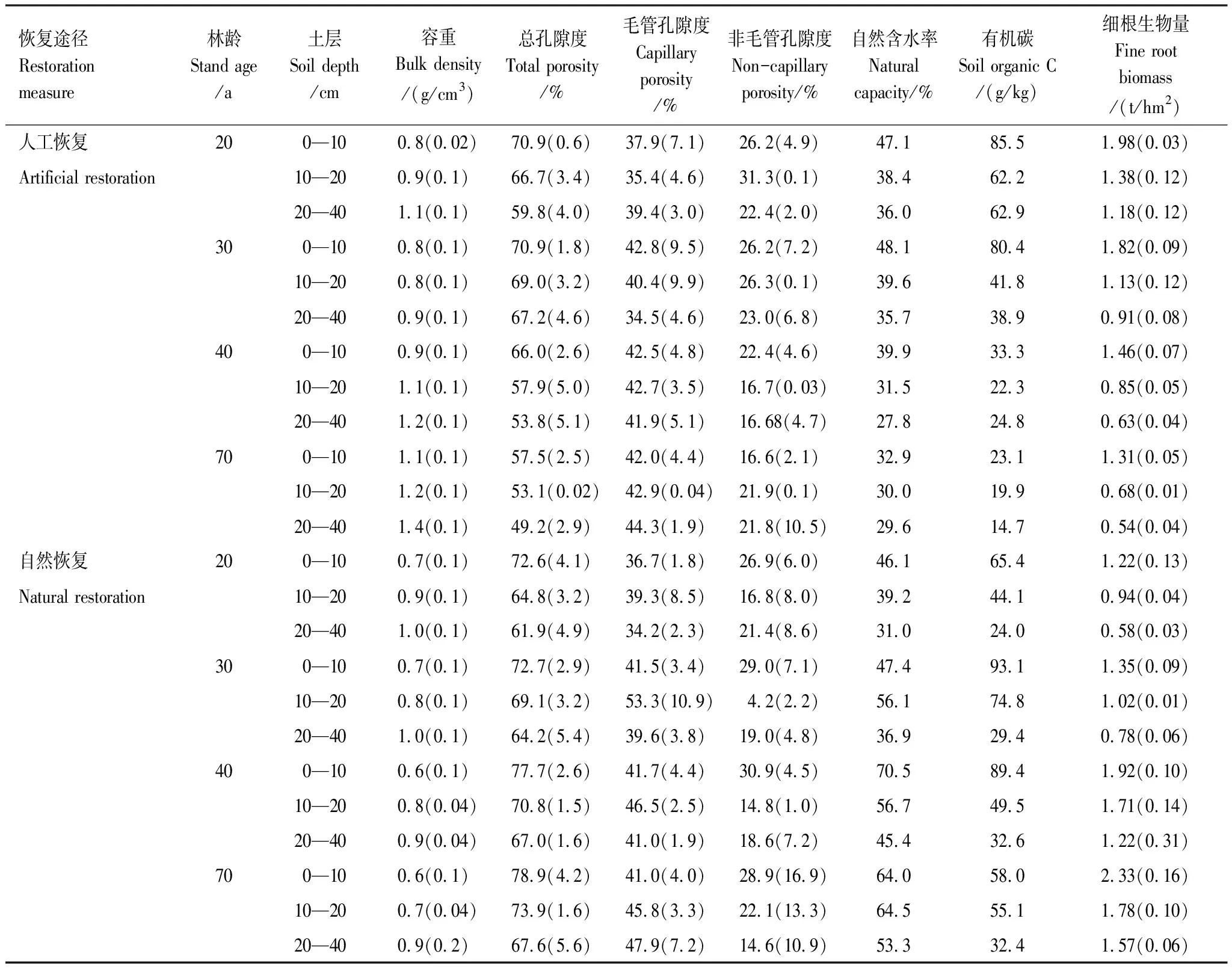
表2 不同恢复途径间土壤理化性质及细根生物量
括号内为标准差(Standard Deviation)
2.2 土壤持水性能
2.2.1 土壤最大持水量
人工恢复云杉林和次生林随着林龄的增加,土壤各层的最大持水量随深度的变化而呈现不同的趋势(图1)。在0—10 cm土层,随着林龄的增加,人工恢复云杉林和次生林土壤最大持水量均没有明显地变化,人工恢复云杉林的最大持水量变化范围为557—588 t/hm2,次生林为534.19—621.19 t/hm2;各恢复途径间0—10 cm土壤最大持水量也没有明显地差异(P>0.05)。同龄段人工恢复云杉林和次生林土壤最大持水量变化率在-8.32%—4.58%间呈现出波动变化,在20—30 a阶段上升,30—40 a阶段下降,40—70 a阶段又上升(图1)。在10—20 cm土层,不同恢复途径间土壤最大持水量的差异显著,在林龄为30—70 a阶段次生林的最大持水量均显著大于人工恢复云杉林(P<0.05)。随着林龄的增加人工恢复云杉林(510—543 t/hm2)最大持水量的变化不显著,自然恢复途径下最大持水量(574—668 t/hm2)显著增大。随林龄的增加,不同恢复途径间土壤最大持水量变化速率在林龄为20—40 a阶段由-5.37%下降至-23.65%,之后又开始上升至-14.17%(图1)。在20—40 cm土层,不同恢复途径间土壤最大持水量存在差异,在林龄为40 a、70 a时次生林的最大持水量均显著大于人工恢复(P<0.05)。随着林龄的增加人工恢复云杉林的最大持水量(1047—1084 t/hm2)变化不显著,次生林的最大持水量(984—1277 t/hm2)均有显著提高。最大持水量变化速率在20—40 a阶段由10.18%迅速下降至-17.20%,之后又开始上升至-8.45%(图1)。总体上,0—40 cm土层最大持水量恢复途径存在明显差异,人工恢复云杉林随着林龄的增加土壤最大持水量(2138—2200 t/hm2)没有明显变化,而次生林随着林龄的增加土壤最大持水量(2142—2565 t/hm2)也在增加,特别在林龄为40、70 a阶段次生林土壤最大持水量要显著大于人工恢复云杉林(P<0.05)(图1)。
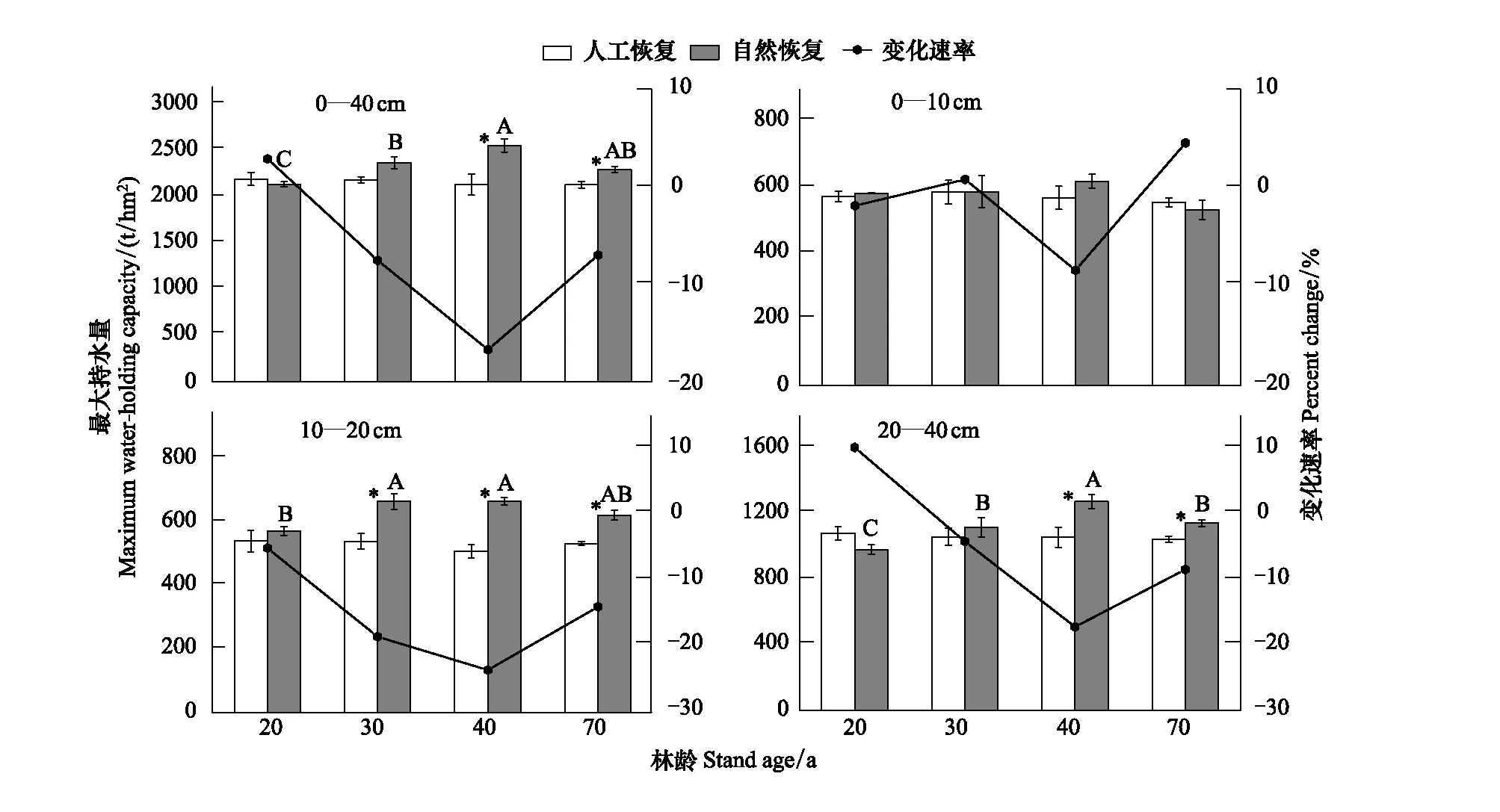
图1 不同恢复途径下各土层最大持水量变化趋势Fig.1 The trends of the maximum water-holding capacity of different soil layers under artificial and secondary restorational chronosequence 不同大写字母代表自然恢复不同林龄间数据差异显著(LSD 检验,P<0.05) ; 竖杠代表标准误;*表示同一林龄下不同恢复途径间差异显著(LSD 检验,P<0.05);变化速率和CNR分别指人工林和次生林土壤最大持水量
2.2.2 土壤毛管持水量
人工恢复云杉林和次生林随着林龄的增加,0—10 cm和10—20 cm土层毛管持水量均没有明显地变化,各恢复途径间土壤毛管持水量也没有明显地差异(P>0.05)(图2)。随着林龄的增加,人工恢复云杉林土壤0—10 cm层毛管持水量变化范围为383—443 t/hm2,次生林变化范围为367—429 t/hm2,人工林与次生林比较,0—10 cm土层毛管持水量变化速率在-3.97%—4.05%间呈现出波动变化。随着林龄的增加,10—20 cm土层毛管持水量变化范围为354—417 t/hm2,次生林的毛管持水量变化范围为427—533 t/hm2,人工林与次生林比较,10—20 cm土层毛管持水量变化速率(-40.00%—-33.91%)呈现出一种平缓的上升状态。随着林龄的增加,人工恢复云杉林和次生林土壤20—40 cm层毛管持水量均显著增大(P<0.05),人工恢复云杉林的毛管持水量变化范围为694—895 t/hm2,次生林为679—942 t/hm2,人工林与次生林比较,20—40 cm土层毛管持水量变化速率在-14.58%—12.51%呈现出波动变化。总体上,随着林龄的增加,人工恢复云杉林和次生林0—40 cm土层毛管持水量逐渐显著升高,人工恢复云杉林土壤毛管持水总量变化范围为1496—1755 t/hm2,次生林为1473—1835 t/hm2,次生林持水量在30林龄阶段显著高于人工云杉林。
2.2.3 土壤非毛管持水量
人工恢复云杉林和次生林随着林龄的增加,各土层土壤非毛管持水量呈现不同的变化趋势(图3)。在0—10 cm和10—20 cm土层,人工恢复云杉林土壤非毛管持水量随着林龄的增加而减小,而自然恢复次生林在0—10 cm土层没有明显地变化,在10—20 cm层70林龄则明显低于其他各演替阶段(P<0.05)(图3),人工恢复云杉林的10—20 cm土层非毛管持水量变化范围为128—191 t/hm2,次生林为136—218 t/hm2,与次生林比较,除40林龄人工云杉林明显较低外,其他各演替阶段变化速率差异不明显。在20—40 cm土层,人工恢复云杉林和次生林随着林龄的增加,土壤非毛管持水量没有明显地变化趋势,与次生林比较,除40林龄人工云杉林明显较低外,其他各演替阶段变化速率差异不明显。总体上,随着林龄的增加,人工恢复云杉林的非毛管持水量(384—702 t/hm2)在显著降低,次生林土壤非毛管持水量(467—793 t/hm2)在40林龄明显大于其他各演替阶段(P<0.05),同时此阶段也是明显大于同时期人工恢复云杉林。
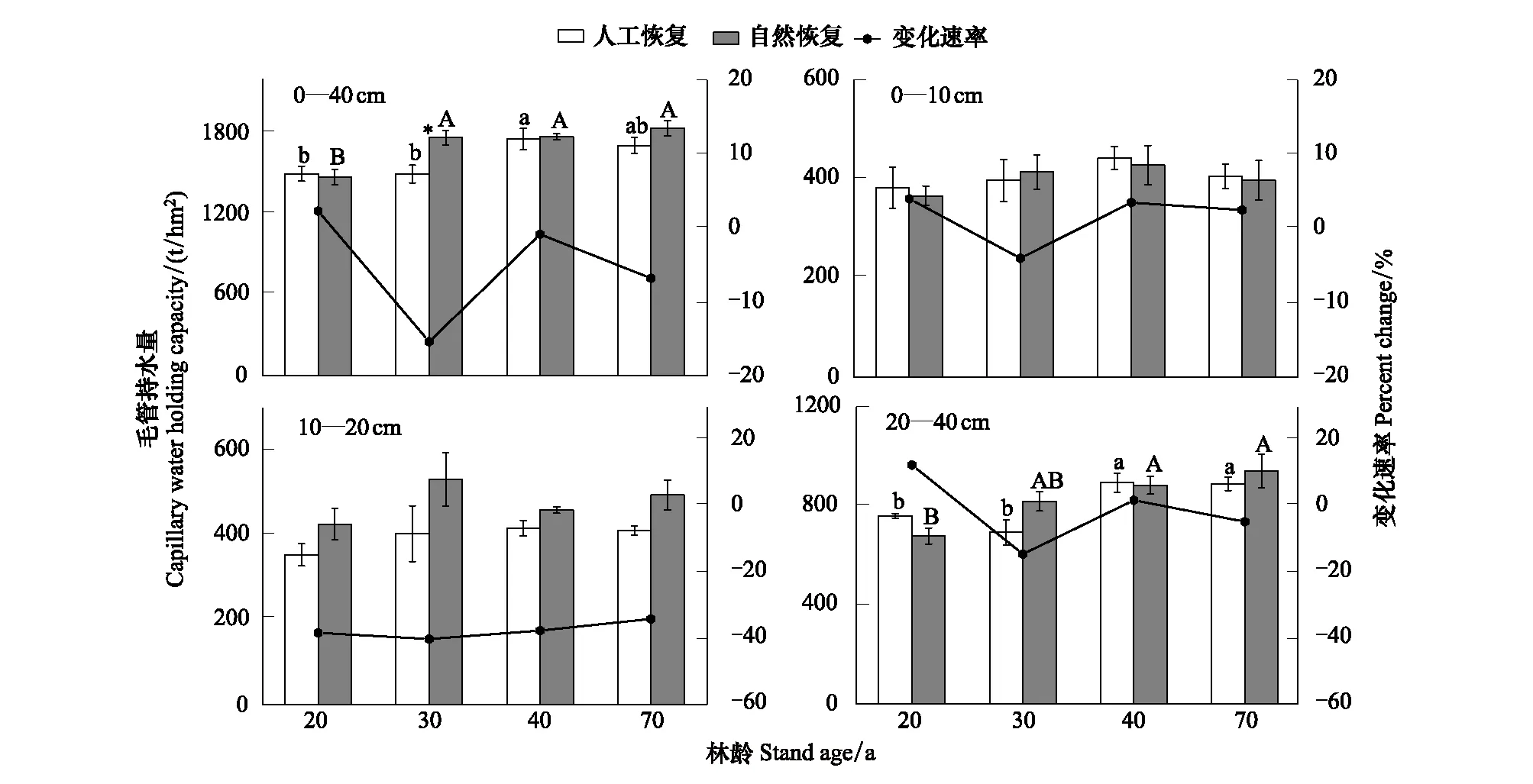
图2 不同恢复途径下各土层毛管持水量变化趋势Fig.2 The trend of the capillary water-holding capacity of different soil layers under artificial and secondary restorational chronosequence不同小写字母代表人工恢复不同林龄间数据差异显著(LSD 检验,P<0.05) ; 竖杠代表标准误;不同大写字母代表自然恢复不同林龄间数据差异显著(LSD 检验,P<0.05) ; 竖杠代表标准误;*表示同一林龄下不同恢复途径间差异显著(LSD 检验,P<0.05);变化速率和CNR分别指人工林和次生林土壤毛管持水量
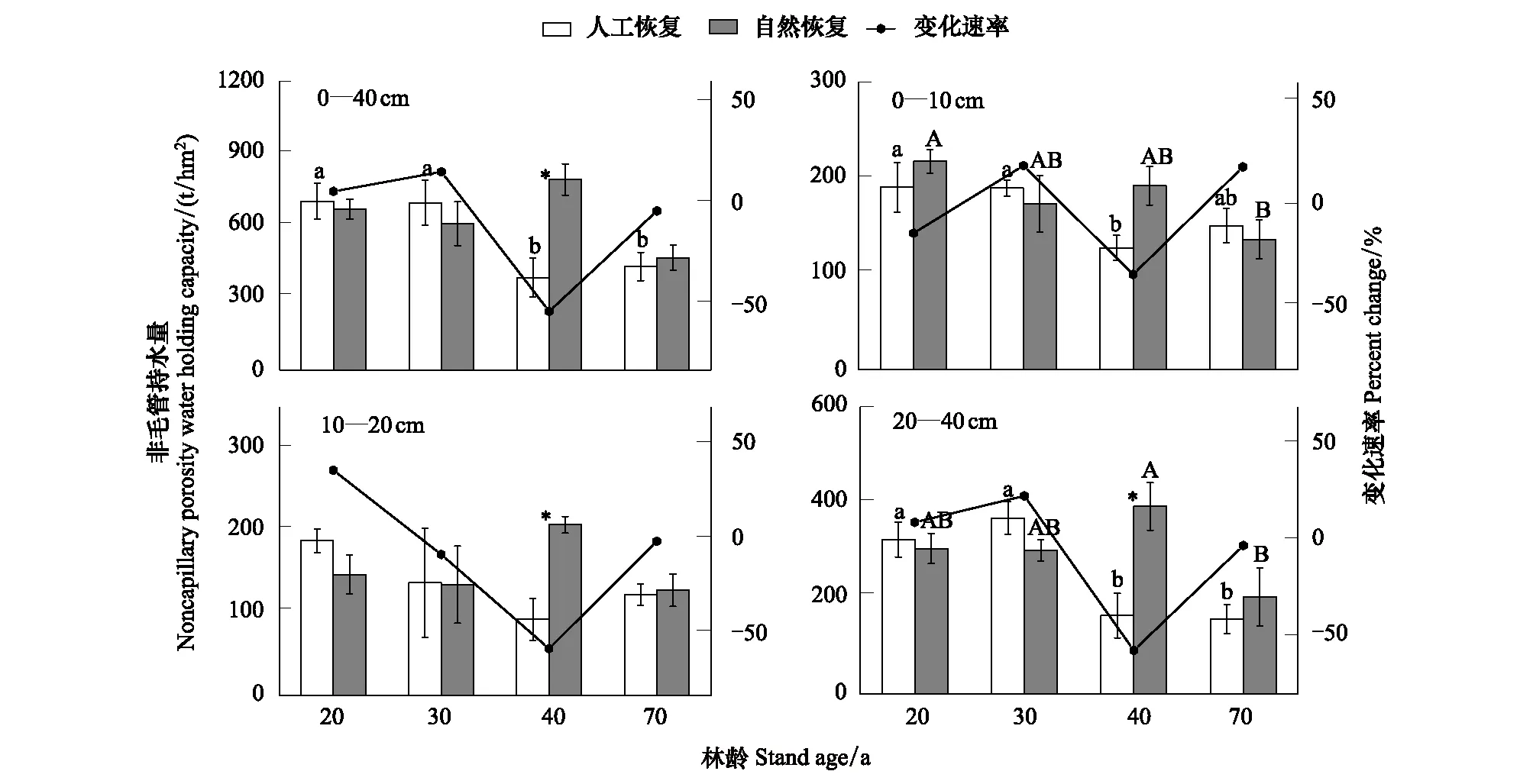
图3 不同恢复途径下各土层非毛管持水量变化趋势Fig.3 The trend of the non-capillary water-holding capacity of different soil layers under artificial and secondary restorational chronosequence 不同小写字母代表人工恢复不同林龄间数据差异显著(LSD 检验,P<0.05) ; 竖杠代表标准误;不同大写字母代表自然恢复不同林龄间数据差异显著(LSD 检验,P<0.05) ; 竖杠代表标准误;*表示同一林龄下不同恢复途径间差异显著(LSD 检验,P<0.05);变化速率和CNR分别指人工林和次生林土壤非毛管持水量
2.3 不同恢复途径土壤持水量与凋落物储量、细根生物量、土壤指标的相关性分析
由相关分析结果(表3)表明,人工恢复途径下土壤最大持水量与土壤总孔隙度、自然含水率、土壤有机碳、细根生物量和凋落物储量有显著的正相关(P<0.01),与土壤容重和土壤毛管孔隙度有显著的负相关(P<0.01);而自然恢复途径下最大持水量与土壤总孔隙度、自然含水率和细根生物量呈显著正相关(P<0.01),与土壤容重(P<0.01)和凋落物储量(P<0.05)呈显著负相关。人工恢复途径下土壤的毛管持水量与土壤容重、土壤毛管孔隙度呈显著正相关(P<0.05),与土壤总孔隙度(P<0.05)、土壤有机碳(P<0.01)、细根生物量(P<0.01)、乔木密度(P<0.01)、凋落物储量(P<0.01)呈显著负相关;而自然恢复途径下土壤毛管持水量与毛管孔隙度、自然含水率和细根生物量呈显著正相关(P<0.01),与郁闭度(P<0.05)、凋落物储量(P<0.01)呈显著负相关。人工恢复途径下土壤的非毛管持水量与非毛管孔隙度(P<0.01)、土壤有机碳(P<0.05)、细根生物量(P<0.05)呈显著正相关;而自然恢复途径下的土壤非毛管持水量只与土壤非毛管孔隙度之间呈显著正相关(P<0.01)。
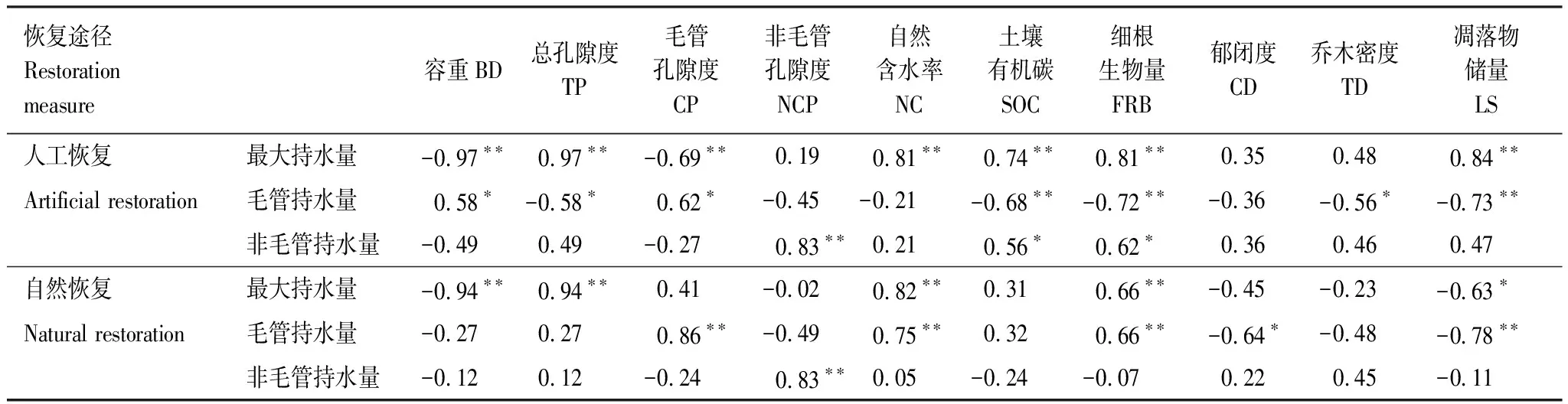
表3 不同恢复途径凋落物储量、细根生物量与土壤指标的相关关系
**P< 0.01,*P<0.05 ;BD: Bulk density; TP: Total porosity; CP: Capillary porosity; NCP: Non-capillary porosity; NC: Nature capacity; SOC:Soil organic carbon; FRB:Fine roots biomass; CD:Canopy density; TD:Tree density; LB:Litter stock
3 结论与讨论
川西米亚罗林区,以云冷杉为优势树种的原始暗针叶林经大规模采伐以后,进行了以云杉树种为主的人工造林恢复,同时,部分区域也同步进行以乡土物种为主的自然更新,目前,该区域森林生态系统处于自然和人工恢复共同主导的格局[4]。相关研究表明,随着植物群落演替或恢复进程的推进,群落结构和物种组成发生变化,进而可能改变土壤性质[21-22]。那么在两种恢复途径进程中,林地土壤性质会发生多大程度的变化?特别是该区域强调生态功能以水源涵养为重点,因此,评估不同恢复途径下土壤涵水能力及其影响因素,对于人工林生态功能提升和调控措施制定具有实践价值和科学意义。本文从林地土壤水源涵养等方面进行了研究,得出了以下结论。
3.1 不同恢复途径对土壤容重及孔隙度的影响
土壤容重及孔隙度等是土壤物理结构的基本指标,能反映土壤涵水能力。在采伐迹地上,通过近70年的人工恢复和自然恢复,部分土壤物理结构和持水保水性能发生了明显改变,但改变的程度和方向由于恢复途径的差异而不同。随着林龄的增加,自然恢复途径下土壤容重及孔隙度相较于人工恢复得到明显改善(表2),可从以下方面进行解释:首先,不同恢复途径由于优势植物组成的不同,引起凋落物分解特性差异,进而导致物质循环与周转速度不尽相同,引起地被物层凋落物蓄积量差异显著[23- 25]。以针叶林为主的人工恢复系列林地的凋落物现存量远大于以阔叶树种为主的自然恢复林地(表1),说明人工恢复林地导致地上凋落物滞留难以归还,而地上凋落物归还是土壤有机质的重要来源,有机质又是改善土壤结构的重要物质基础[7]。同时,凋落物回归速率受阻,将导致以凋落物为载体的C、N、P元素的归还速率减缓[26],进而影响有机质对土壤物理性质的改良效果[27-28]。其次,与自然恢复次生林相比,人工恢复云杉林中林下修枝等森林经营的人为动干扰增加了土壤紧实度[29]。另外,林型结构差异可能也是引起林内土壤物理性质差异的重要原因[30]。由于人工恢复云杉林的造林树种单一,种植密度大(表1),郁闭度大于自然恢复次生林,阻挡了大量的光照与降水,阻碍了林下植被的生长与更新,加之其凋落物分解速度慢,无法及时归还养分改善土壤的物理结构,随着林龄的增加,使得人工恢复相对于自然恢复样地,土壤容重增加,孔隙度降低(表2)。最后,根系生物量及分布规律也对土壤容重和孔隙度产生了影响[31]。一方面,土壤有机质的来源除了地上凋落物,地下细根根系周转和分泌物等也贡献了相当一部分[32-33],随着林龄的增加自然恢复林地的细根生物量要大于人工云杉林地(表2)。来源于凋落物及根系的有机质分解形成腐殖质,同根系残体及其分泌物胶结联接土壤颗粒[28],促进孔隙形成[34]。另一方面,根系的穿插可增加土壤空隙,特别是毛管孔隙[35],由于表层土中根系分布较多,因此经过近70 a的恢复后,孔隙的改善也主要集中在表土层(表2)。林地土壤上层分布密集的根系通过在土壤中穿插、挤压,促进了土壤发育,改善了土壤结构及孔隙状况,因此表层孔隙度显著大于下层[35]。随着林龄的增加自然恢复的土壤孔隙度均大于人工恢复样地,可从各土层的根系生物量明显大于人工恢复林地来部分解释(表2)。
3.2 不同恢复途径对土壤持水性的影响
土壤水分特征除了受区域气候特征等外界条件影响外,植被类型也是一个重要的影响因素[36-37]。植被类型的差异通过改变林下微环境、凋落物回归、有机质含量和根系分布及数量等来影响土壤结构[38-39]。如上关于恢复途径差异对土壤物理结构的影响因素所述,自然恢复林地的树种以阔叶树种为主,相较于人工恢复的针叶树种更易于分解,使得其地表覆盖大量枯枝落叶层分解增加了土壤有机质,同时,根系的穿插也使得土壤孔隙度显著增加,而根系死亡留下的空隙和根系周围的生物的活动都增加了土壤的非毛管孔隙,促使土壤形成良好结构,使土壤孔隙度和通气状况变好,有利于水分的保持和养分的传输与运送[35]。这些因素通过改变土壤物理结构而间接影响土壤持水特性。
土壤物理结构及孔隙特性是土壤涵水能力的基础,即土壤容重和孔隙度的变化直接影响土壤持水量,进而改变土壤的通气、蓄水和持水特性[40]。本研究表明随着林龄的增加自然恢复林地的最大持水量要大于人工恢复林地(图1),通过相关性分析结果(表3)可知,两种恢复途径的最大持水量都与土壤容重呈显著负相关,与土壤总孔隙度、细根生物量呈正相关。与此对照,自然恢复林地的最大持水量与凋落物储量呈显著负相关(-0.63,P<0.05),人工恢复则成显著正相关(0.84,P<0.01),说明两种恢复途径凋落物归还差异可能是导致土壤最大持水量差异的主要原因,而土壤有机碳含量在人工林中与土壤最大持水量呈显著正相关(0.74,P<0.01),而在自然恢复途径中二者没有明显相关性,说明在人工恢复途径下,有机质归还对于土壤最大持水量改善起重要作用。同时,随着林龄的增加两种恢复途径0—40 cm土壤毛管持水量也显著增加,但自然恢复的土壤毛管持水量显著大于人工恢复(图2),通关相关性分析可知,两种恢复途径的毛管持水量都与毛管孔隙度呈现出显著正相关,和凋落物储量呈现出显著负相关。但细根生物量对两种恢复途径下土壤毛管持水量有不同的影响(表3),即在人工恢复途径下土壤毛管持水量与细根生物量呈明显负相关(-0.72,P<0.01),而在自然恢复途径下土壤毛管持水量与细根生物量呈明显正相关(0.66,P<0.01),说明细根是导致两种恢复途径下土壤毛管持水量差异的另一原因。
总体而言,通过近70 a的恢复,由于恢复途径的不同导致了土壤水源涵养能力的差异。随着恢复年限的增加,自然恢复途径下次生林土壤容重降低,孔隙度持续增加,持水能力优于人工恢复云杉林。由于人工恢复的云杉林凋落物质地特性的差异[41],导致凋落物分解慢,有机质回归受阻[27-28]。自然恢复以桦木为主的混交林土壤SOC (表2)含量的增加得益于其凋落物的易分解性,一方面改善了土壤结构,使土壤松软多孔;另一方面,改变了土壤的胶体状况,使土壤吸附作用增强。这些土壤结构特性的改变都有利于改善土壤通气、持水性和细根的穿插与发育,反之,凋落物分解缓慢致使有机物回归土壤受阻,间接影响土壤物理结构,导致林地土壤的水土保持能力弱。通过进一步的相关性分析表明,两种恢复途径下凋落物归还、有机碳含量和细根可能是影响林地土壤物理结构和持水能力的主要因素。这些结果说明,在对采伐迹地或退化生态系统进行造林恢复时,应尽量避免营造结构单一和高密度的人工纯林,同时,在对现有人工纯林进行功能提升改造时,应借助乔木结构调控的方法,改造结构单一的人工纯林为结构复杂的复层近自然林,为物质循环、土壤生态功能提升提供条件。
