松嫩平原盐碱草地土壤酶活性与植物群落特征的关系初探
2018-12-20,,,,
,,,,
(哈尔滨师范大学生命科学与技术学院,黑龙江省普通高等学校植物生物学重点实验室,黑龙江 哈尔滨 150025)
松嫩平原草地是我国著名的天然草场,又是东北西部的绿色生态屏障,也是我国重要的畜牧业基地之一,具有较高的经济价值和重要的生态意义。但随着自然环境的变化以及人类活动的干扰,草地盐碱化程度逐渐加重,盐碱化的加剧会对土壤微生物活性和群落组成产生直接的影响[1-2],也会通过影响植物根系和土壤动物的活动而对土壤微生物群落产生间接的作用,最终影响土壤酶的活性[3-5]。土壤中的一切生物化学反应,实际上都是在酶的参与下进行的,土壤的酶活性反映了土壤中进行的各种生物化学过程的强度和方向,它是土壤的本质之一[6],也是土壤变化的敏感指标[7]。Dick等[8]早在1992年就指出:“土壤酶有助于描述和预测不同生态系统的功能、质量及各系统间的相互作用”。进入21世纪以后,不同生态系统土壤酶活性的研究已经成为生态学的热点问题之一。在草地生态系统中,人们对土壤酶的研究主要集中在以下几个方面:1)不同类型草地土壤酶活性研究,涉及的草地类型包括草甸草地[9]、荒漠草地[10-11]、高寒草地[12-13]、盐碱草地[14-16]、沙化草地[17]等;2)草地逆行演替不同阶段土壤酶活性研究,包括轻度、中度、重度退化草地[18-21];3)人为干扰对草地酶活性影响的研究,如放牧[22-23]、刈割[24-25]、施肥[26]、外来污染[27-30]等;4)不同恢复措施下草地土壤酶活性研究,包括合理放牧[31]、围封禁牧[32-33]、土耕恢复[34]等。从这些研究不难看出,学者们关注的焦点在于不同生物、非生物环境对土壤酶活性的影响,而在草地土壤-植被连续系统中,地上生物环境变化(植物群落)是否会引起土壤酶活性的响应还较少涉及,因此在前期研究工作的基础上[35-36],进一步探讨松嫩平原盐碱草地土壤酶活性与植物群落特征的关系,为盐碱草地的合理利用和改良提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区域位于松嫩平原的西南部草地,地理位置为45°53′-47°08′ N和123°45′-124°42′ E,平均海拔152 m。属温带大陆性季风气候,冬长雪少,天气寒冷;夏短湿热,降水集中;春季风大,气候干燥;秋凉气爽,时有早霜。年平均气温4.0 ℃,月平均气温稳定在0 ℃以上的日数为210 d,年降水量300~500 mm。土壤类型为草甸盐碱土,一般盐分含量为15~100 g·kg-1。
1.2 样品采集与处理
2016年5-9月的每月15-20日,5月平均气温11.5 ℃,降水总量36 mm;6月平均气温18.5 ℃,降水总量74 mm;7月平均气温21 ℃,降水总量127 mm;8月平均气温19 ℃,降水总量110 mm;9月平均气温12 ℃,降水总量61 mm。在研究区域内随机选取1 m×1 m的典型样方9个,在每个样方内进行植物群落调查,记录植物种类、盖度、高度等群落特征。5月样地内植物有8科15属13种、6月样地内植物有12科22属22种、7月样地内植物有9科19属20种、8月样地内植物有10科22属22种、9月样地内植物有8科16属18种。主要的植物有羊草(Aneurolepidiumchinense)、苔草(Carextristachya)、虎尾草(Chlorisvirgata)、鹅绒委陵菜(Potentillaanserina)、蔓委陵菜(Potentillachinensis)、蒲公英(Taraxacumofficinale)、车前草(Plantagoasiatica)、黄蒿(Artemisiascoparia)、稗草(Echinochloacrusgalli)等。
将所有地上植被沿地表剪下,按照植物种类分别封装。然后在每个样方内去除植被残体,随机选取3个样点,采集0~20 cm土层土壤样品,充分混合。将植物和土壤样品带回实验室,将植物样品烘干,用于生物量的测定;将土壤样品除去植物残根,自然风干后,过1 mm筛,用于土壤酶活性的测定。
1.3 土壤酶活性的测定方法
过氧化氢酶(catalase,CAT)活性的测定采用高锰酸钾滴定法,多酚氧化酶(polyphenol oxidase,PPO)活性和过氧化物酶(peroxidase,POD)活性的测定采用邻苯三酚比色法,蛋白酶(protease,PRO)活性的测定采用茚三酮比色法,碱性磷酸酶(alkline phosphatase,AKP)活性的测定采用磷酸苯二钠比色法,转化酶(invertase,INV)活性的测定采用二硝基水杨酸比色法[37]。
1.4 植物群落特征计算方法
利用Pielou均匀度指数、Shannon-Wiener多样性指数、Simpson多样性指数、丰富度指数、物种均匀度指数来表征各样地的植物群落特征。各指数的计算方法如下:
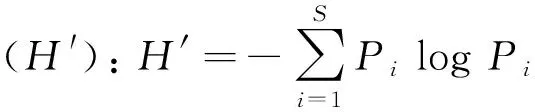

丰富度指数(R):R=S
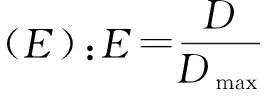
式中:S为物种总数;ni为抽样中第i个物种的重要值;N为抽样中所有物种的重要值总和;Pi是第i个物种的重要值ni占所有种的重要值总和N的比例,即Pi=ni/N。
重要值=[(相对重量+相对盖度+相对密度+相对高度)/4]×100%
1.5 数据分析方法
采用SAS 9.2软件对试验数据进行单因素方差分析、相关分析和逐步回归分析;应用典范对应分析(canonical correspondence analysis, CCA)对土壤酶活性与植物群落特征的关系进行分析,通过CANOCO 4.5软件建立二维排序图。采用Excel软件作图。
2 结果与分析
2.1 盐碱草地生长季内土壤酶活性的变化
在植物不同生长季内,盐碱草地6种土壤酶活性的变化规律不同(图1)。土壤多酚氧化酶和过氧化物酶活性在5月较高,随后降低,到8月最低,9月又升高(图1A,图1C);土壤过氧化氢酶活性、转化酶活性先升高后降低,峰值出现在8月(图1B,图1F);土壤蛋白酶活性先降低后升高,最低值出现在7月(图1D);土壤碱性磷酸酶活性在6月降低,随后升高,到9月达到最大(图1E)。
2.2 盐碱草地生长季内植物群落特征的变化
在植物不同生长季内,盐碱草地植物群落特征的变化规律不同(图2)。Pielou指数在整个生育期变化较小(图2A);Shannon指数和丰富度呈“双峰”变化,在6和8月较高,而在5、7、9月较低(图2B,图2D);Simpson指数在8月最高,其他月份较低(图2C);均匀度指数先降低后升高,在6月最低,而在9月最高(图2E);生物量先增加后降低,峰值出现在8月(图2F)。
2.3 盐碱草地生长季内土壤酶活性与植物群落特征的关系
2.3.1土壤酶活性与植物群落特征的相关分析 对整个植物生长季内的土壤酶活性和植物群落特征关系进行相关分析(表 1),结果表明多酚氧化酶活性和Shannon 指数显著负相关(P<0.05);过氧化物酶活性和Shannon指数、丰富度指数、Simpson指数显著负相关(P<0.01,P<0.05);转化酶、碱性磷酸酶活性和Pielou指数显著正相关(P<0.05);过氧化氢酶、蛋白酶活性和植物群落特征间无显著相关关系(P>0.05);植物群落特征中的均匀度指数和生物量与酶活性间没有显著的相关关系(P>0.05)。

图1 不同生长季内土壤酶活性的变化Fig.1 Changes of soil enzyme activities in different growing season
2.3.2土壤酶活性与植物群落特征的逐步回归分析 对具有相关关系的多酚氧化酶、过氧化物酶、碱性磷酸酶、转化酶和Pielou指数、Shannon 指数、Simpson指数、丰富度指数进一步进行回归分析,结果表明,多酚氧化酶活性在8月与Simpson指数有较好的量化关系,关系式为Y=-0.009+0.053X(F=5.72,P<0.05);过氧化物酶活性在7月与Shannon 指数有较好的量化关系,关系式为Y=2.047-1.950X(F=7.56,P<0.05);碱性磷酸酶活性在9月与Pielou均匀度指数有较好的量化关系,关系式为Y=1.535+4.388X(F=9.31,P<0.05);转化酶活性在7月与丰富度指数有较好的量化关系,关系式为Y=2.887+0.551X(F=6.11,P<0.05)。
2.3.3土壤酶活性与植物群落特征的典范对应分析 在整个生长季内,对不同月份土壤酶活性和植物群落特征进行典范对应分析(图3,表2),结果表明,在研究的6种土壤酶中,只有过氧化物酶活性出现在第一排序轴,其特征值为0.107,梯度长度为0.523,其余5种土壤酶活性均未出现在图中,说明多酚氧化酶、过氧化氢酶、蛋白酶、碱性磷酸酶和转化酶活性与植物群落特征没有典范对应关系。过氧化物酶活性与各植物群落特征都是负对应关系,对应大小的顺序是:丰富度指数>Shannon多样性指数>均匀度指数>Simpson多样性指数>生物量>Pielou均匀度指数。
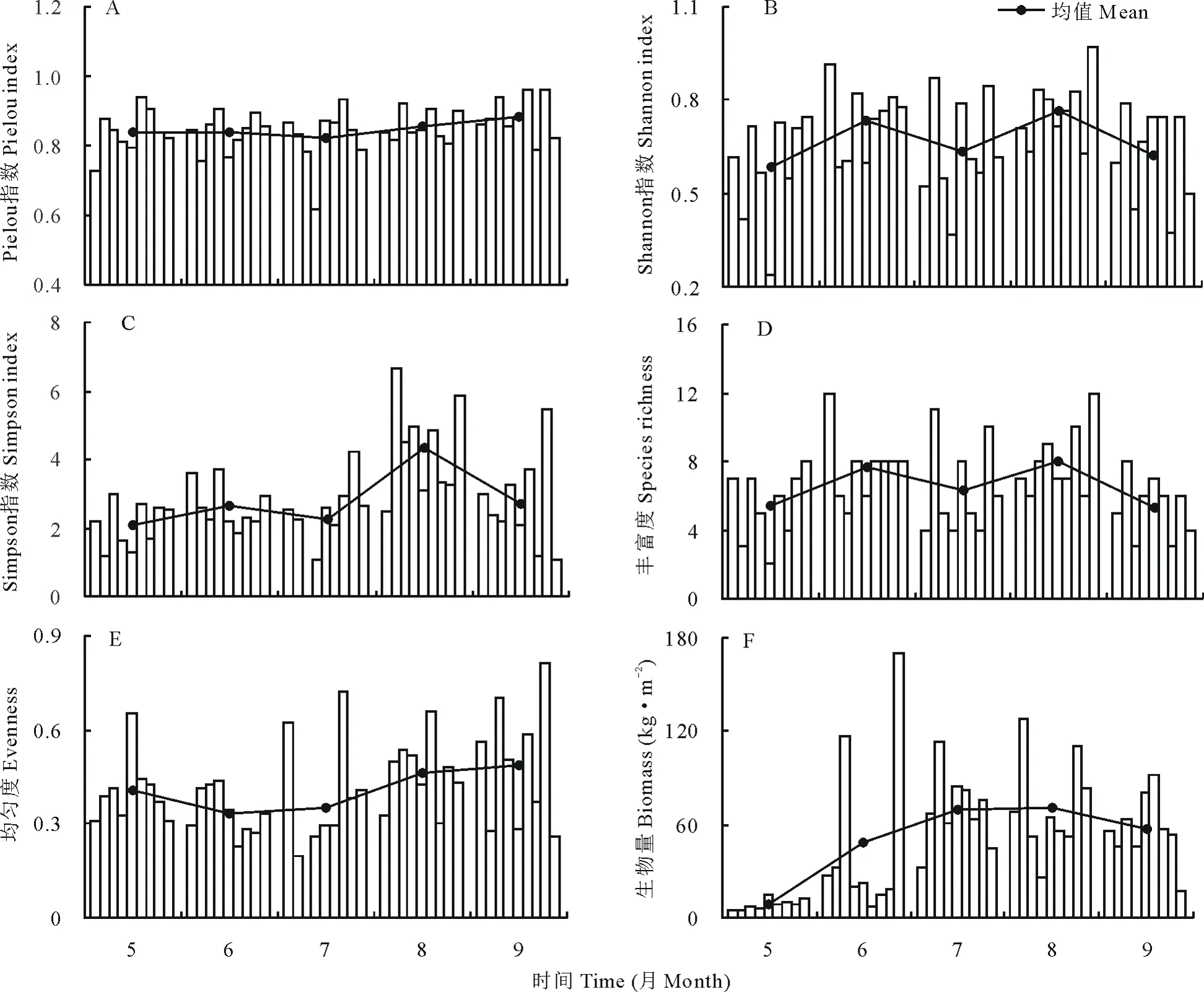
图2 不同生长季内植物群落特征的变化Fig.2 Changes of plant community characteristics in different growing season
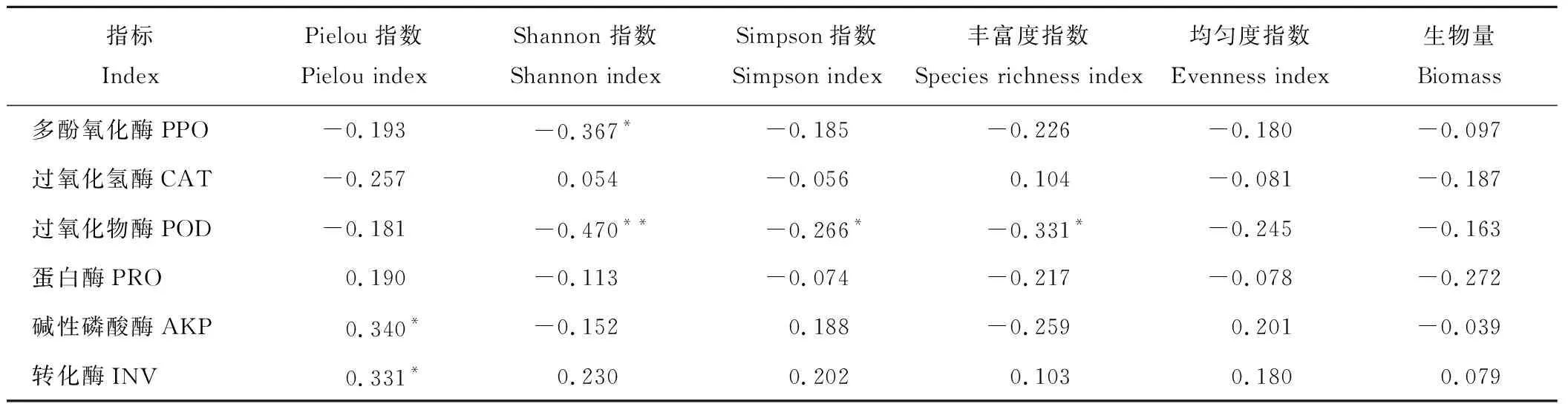
指标IndexPielou指数Pielou indexShannon 指数Shannon indexSimpson指数Simpson index丰富度指数Species richness index均匀度指数Evenness index生物量Biomass多酚氧化酶PPO-0.193-0.367*-0.185-0.226-0.180-0.097过氧化氢酶CAT-0.2570.054-0.0560.104-0.081-0.187过氧化物酶POD-0.181-0.470**-0.266*-0.331*-0.245-0.163蛋白酶PRO0.190-0.113-0.074-0.217-0.078-0.272碱性磷酸酶AKP0.340*-0.1520.188-0.2590.201-0.039转化酶INV0.331*0.2300.2020.1030.1800.079
注:* 表示相关显著(P<0.05),** 表示相关极显著(P<0.01);n=45。
Note:* indicate significant correlation on the 0.05 level, ** indicate significant correlation on the 0.01 level;n=45.
3 讨论

图3 土壤酶活性与植物群落特征间的典范对应分析Fig.3 The CCA analysis of soil enzyme activity and plant community characteristics A: 均匀度指数Evenness index; B: Simpson多样性指数Simpson index; C: Pielou均匀度指数Pielou index; D: 生物量Biomass; E: Shannon多样性指数Shannon index; F: 丰富度指数Species richness index;图中数字代表不同月份Numerals represent different months.
3.1 盐碱草地生长季内土壤酶活性的变化
在以往草地研究中,学者们研究得比较多的酶类有过氧化氢酶、磷酸酶、转化酶、脲酶等[38-40],这些酶类在土壤呼吸及土壤碳、氮、磷循环中具有重要作用,而课题组在研究中发现多酚氧化酶、过氧化物酶与土壤代谢间也存在着密切关系[16,18,41],因此选择了多酚氧化酶、过氧化氢酶、过氧化物酶、蛋白酶、碱性磷酸酶和转化酶这6种典型酶类来进行研究。研究发现,随着生育期的变化,各个酶表现出一定的顺序作用,在春季(5、6月)多酚氧化酶、过氧化物酶、蛋白酶和碱性磷酸酶的活性都较高,说明土壤代谢过程比较活跃,土壤氮、磷代谢旺盛,到夏季时(7、8月)过氧化氢酶、转化酶活性升高,土壤呼吸和土壤碳代谢强烈,而9月各个酶恢复到原来水平,这与朱丽等[42]、李凤霞等[43]研究的过氧化氢酶、转化酶、多酚氧化酶及碱性磷酸酶活性的变化规律一致。过氧化物酶在松嫩盐碱草地中的研究还较少报道,需要在以后的工作中进一步研究探讨。
3.2 盐碱草地生长季内植物群落特征的变化
植物群落的生产力和物种多样性是表征群落结构和功能的重要量化指标[44],它们是植物生物学特性和外界环境条件共同作用的产物,也可以反映不同自然地理条件与群落的相互关系[45]。Shannon多样性指数能客观地反映物种的丰富度和均匀度,Pielou均匀度指数能较好地反映物种的均匀程度,Simpson指数能较好地反映群落的优势度,丰富度指数能较好地反映不同植物群落类型在物种组成方面的差异,物种均匀度是指一个群落或环境中的全部物种个体数目的分配状况。本研究中,不同盐碱草地植物群落特征随季节变化呈现不同的变化规律,这是由于盐碱草地的异质性较强,不同样地植被多样性变化较大,而不同植被的生长发育特性不同,在不同生育期对环境的适应能力也不同,只有生物量的变化与以往研究的单一群落如碱蓬(Suaedaglauca)群落、虎尾草群落、羊草群落生物量的季节动态变化有相似之处[46],但Pielou均匀度指数、Shannon多样性指数、Simpson指数、丰富度指数、均匀度指数等群落特征指数的变化还无一定的规律可循,在研究时要综合考虑气候及土壤环境因子的变化进行探讨。

表2 植物群落分布DCA排序Table 2 DCA ordinations of plant communities
3.3 盐碱草地生长季内土壤酶活性与植物群落特征的关系
在土壤-植物-大气连续体(soil-plant-atmosphere continuum,SPAC)中,土壤、植被生长以及外界环境间会存在某种联系,在不同生态系统中表现也会不同[47]。在草地生态系统中,学者们很少关注地上植被生长与土壤特性间的关系,仅有的研究表明,松嫩盐碱草地植物群落特征受土壤养分、盐分的共同影响,在不同季节各种土壤因子和植物群落特征的关系不同[48]。尽管研究中发现土壤酶(多酚氧化酶、过氧化物酶、碱性磷酸酶、转化酶)和植物群落特征(Pielou指数、Shannon 指数、Simpson指数、丰富度指数)间存在一定的相关关系或在某个月份表现出量化关系,但将土壤酶活性与植物群落特征作为两组变量进行CCA分析后,这种对应关系基本消失(过氧化物酶除外),说明在SPAC系统中,还很难将土壤酶的活性和植物群落特征指数联系在一起,这可能是因为地上植物的生长并不会立即对土壤生物产生直接的影响,尤其当植物群落特征分异较大时,植物群落特征分异与土壤生物学的变化并不同步,植物群落特征发生反应需要经过一定的时间,其生长后的凋落物进入土壤后才会引起土壤生物发生响应。至于在何种生境中植物群落特征会与土壤酶存在某种联系,还有待于进一步研究。
值得一提的是,过氧化物酶活性与植物群落特征间存在较好的对应关系,过氧化物酶是土壤中一种重要的氧化还原酶类,能利用微生物活动和某些氧化酶(例如尿酸盐氧化酶)的作用来氧化土壤有机物质,在腐殖质的形成中具有重要作用[49]。本研究发现过氧化物酶表现为与各群落特征指数呈负相关,这说明在盐碱草地盐碱化过程中,过氧化物酶活性逐渐增强,这可能是土壤对不良环境的一种适应性反应,有利于土壤在逆境条件下进行有机质的代谢过程,可以在盐碱草地研究中,尝试将过氧化物酶作为一个敏感的指标来指示植物群落的变化过程。但需要指出的是研究过氧化物酶活性在盐碱草地中的变化规律只是对土壤生物学指标的一种初步尝试,还应选择不同的酶类及不同区域的盐碱草地类型进行深入研究。
4 结论
1)在不同生长季内,土壤酶活性和植物群落特征变化规律不同,与以往研究结论一致。2)在整个生长季内,多酚氧化酶活性和Shannon 指数显著负相关(P<0.05);过氧化物酶活性和Shannon指数、Simpson指数和丰富度指数显著负相关(P<0.01,P<0.05);转化酶、碱性磷酸酶活性和Pielou指数显著正相关(P<0.05)。3)在不同生长季内,土壤酶活性和植物群落特征关系不同。7月过氧化物酶活性与Shannon指数、转化酶活性与丰富度指数有量化关系,8月多酚氧化酶活性与Simpson指数有量化关系,9月碱性磷酸酶活性与Pielou指数有量化关系。4)CCA分析结果表明过氧化物酶与植物群落特征间存在较好的对应关系。
