基于群落结构及土壤理化性质对围封7年青藏高原东南缘高山草地的综合评价
2018-12-20,,,*,,,,,,*
,,,*,,,,,,*
(1.中国科学院成都生物研究所,四川 成都 610041;2.中国科学院大学生命科学学院,北京 101408;3.华东师范大学生态与环境科学学院,上海 200241;4.中国科学院地理科学与资源研究所,北京 100101;5.阿坝师范学院资源与环境学院,四川 阿坝623002)
放牧是人类利用和管理草地的重要方式,在草地生态系统中发挥着物质循环、能量转化和信息传递等重要作用[1-3]。纵观人类发展历程,放牧不仅给人类的生存发展提供了物质基础,而且在文化的交流上起到了积极作用[4]。作为重要的农业生产活动,放牧既是牧民经济收入的来源,同时也为草地生态系统提供了一种保护机制,避免了集中式饲料养殖引发的环境污染及食品安全等问题[5]。据统计,全球牧区面积约占陆地面积的50%以上,主要分布于美国、澳大利亚、新西兰等具有温带大陆性气候或热带草原气候的国家和地区[2];我国牧区面积约占国土面积的41%,主要分布于胡焕庸线的左侧,属于半干旱区域的脆弱生态带[2,6]。放牧作为我国农业结构中的重要组成部分,使广大人民普遍受益,但它同样是把“双刃剑”。随着社会经济的增长和人们物质需求的逐步提高,过牧现象大面积出现,导致草地生态系统结构组成发生了变化,其功能亦随之减弱。主要体现在如下几个方面:首先,过度放牧致使植物群落结构的组成明显改变,牲畜的选择性采食使物种多样性单一化,并让毒害杂草成为优势物种[7-8];其次,牲畜活动导致土壤碳、氮等元素含量降低,其践踏作用导致土壤容重增加,土壤质量逐渐下降[9];再者,牲畜数目超过单位面积草地的承载量,打破植被生长的良性循环,使草地生产力降低,草地生态系统的稳定性及其功能遭到严重破坏[10],因此,应积极采取措施防止草地生态系统继续恶化,使其在科学有效的管理方式下得以修复,既而可持续地服务于人类社会。
围栏封育、重建人工草地等是草地修复的主要措施,在我国不同的草地类型中有着广泛应用。围栏封育因其投资小,见效快以及方便管理为世界各地采用,其修复机制主要是通过排除放牧的干扰,使草地生态系统凭借自身弹力得以恢复[11-12]。在实际应用过程中,围栏封育的确取得了良好的生态及经济效益,但一些研究者也对其实用性及修复效果提出了质疑,即是否所有退化草地都可以进行围栏封育,以及关于围封时间与草地修复情况。已有研究表明围栏封育并不具有普适性,不恰当的围封方式及围封时间带来的修复效果并不理想,甚至会加速草地退化[13],苏建红等[14]对高寒草甸的长期围封实验表明围栏内的土壤全氮及全磷含量低于围栏外,仅土壤含水量及容重得到改善。刘建等[15]发现围封19年后草地生物多样性下降,Vickery[16]和Wang等[17]指出围栏破坏了放牧带来的牧草超补偿生长机制,而导致草地生产力降低,更有学者提出围栏封育不仅没有改善草地质量,还严重地破坏了野生动物的迁徙廊道[18-19]。因此,对围栏封育后的草地质量进行综合系统的评估非常必要,有助于明确围栏封育的修复机制及其所带来的实际效益。
作为我国四大牧区之一的青藏高原,具有独特的地理条件与气候因素,对全球变化的响应非常敏感,同时也使得其草地生态系统脆弱化[20-22]。放牧作为青藏高原长期维持生计的活动方式有着悠久的历史。但是,近年来不合理的放牧方式使得草地生态系统岌岌可危,同时学者们也开展了相应的草地修复工作。目前,国内外对草地修复机制及效果方面进行了大量研究,但主要局限于独立指标的研究,如植被高度、生物量、生物多样性等的研究[23-25],此外,亦有针对退化草地及其恢复后土壤理化性质的研究[14-15, 26]。然而,针对独特的低纬度高海拔退化高山草地围封后,通过植物-土壤互作界面进行系统性的评价研究则相对较少。基于此背景,本研究选取青藏高原东缘岷江源区的典型退化高山草地作为研究对象,对比围封7年后高山草地围栏内外的植物生物量、多样性及土壤理化性质,探究其变化规律及响应程度,进而对草地质量进行综合评价,旨在系统地认识退化草地的恢复过程,并为高山草地修复工作提供数据基础与科学依据,进一步阐述其生态学内涵与意义。
1 材料与方法
1.1 研究区概况
本试验地位于四川省松潘县卡卡山山顶(32.59° N, 103.40° E),属于青藏高原东缘的岷山山脉中部,海拔为3850~3950 m,该地区受西南暖湿气流和东南季风控制,属于典型山地季风气候。年均气温6.7 ℃,1月均温-4.3 ℃,7月均温15.6 ℃,年均降水量为756 mm,并呈现明显的季节性分布,超过70%的降水出现在夏季(6-8月)。年均太阳辐射时数1827.5 h,≥ 0 ℃年积温1833.6 ℃,无绝对无霜期。土壤为棕色草毡土,土壤pH 5.61~7.11,有机质29.31~88.28 g·kg-1干土,全氮2.64~7.97 g·kg-1干土,平均土层厚度63 cm,植物根系主要分布于2~20 cm 土层。苔藓类植物主要有微齿金发藓(Polytrichumswartzii)和长蒴藓(Trematodonacutus);维管束植物主要有嵩草属(Kobresia)和薹草属(Carex),其他常见物种还有高原毛茛(Ranunculustanguticus)、甘肃马先蒿(Pediculariskansuensis)、川西小黄菊(Pyrethrumtatsienense)、珠芽蓼(Polygonumviviparum)、盘花垂头菊(Cremanthodiumdiscoideum)和滨发草(Deschampsialittoralis)等[27-28]。
1.2 试验设计及样品采集
2006年9月,在卡卡山山顶选择一片相对平坦的区域,用铁丝网建立60 m×60 m的围栏,其内外分别设置6个小区,大小为10 m×10 m,为避免边际效应,小区距离围栏5 m左右,每个小区间隔5 m。该区域放牧时间大致为9月下旬至次年7月上旬,放牧强度为每hm24~5头牦牛,草地利用率约为75%,属于重度放牧。2012年9月在每个小区内随机选取10个0.25 m×0.25 m小样方,剪取并收集地上植物及其凋落物,分类并记录每个小区内的物种数。收集到的植物在105 ℃杀青30 min后置于65 ℃烘干至恒重(48 h)并称量。围栏内外每个小区内于0~10 cm层次各挖取土壤样品5份,剔除根系和杂物,带回实验室混匀后用于土壤理化性质测定;2012年6月分别于任意3个小区内设置一个纽扣式温度计(iButton-TMEX RTE), 埋置于地表下15 cm处,测量2012年6月至9月的土壤温度,每小时测量一次。另外用环刀在每个小区内随机取土壤样品2份,分别用于土壤含水量与土壤容重的测定。
1.3 样品分析
不同层次挖取的每份土壤样品分别混匀过筛(2 mm)测定理化性质。采用氯仿熏蒸法测定土壤微生物生物量碳、氮含量;采用重铬酸钾氧化(外加热)法测定土壤有机碳含量;采用凯氏法测定土壤全氮含量;采用紫外分光光度法测定硝态氮及铵态氮含量;采用钒钼黄比色法测定土壤全磷含量;采用火焰分光光度法测定全钾含量;采用乙酸钠-火焰光度法测定阳离子交换量;采用电导法测定土壤pH值(水土比为2.5∶1.0);采用烘干法测定土壤含水量及容重[29]。
群落多样性指数按照下列公式测定[30]:

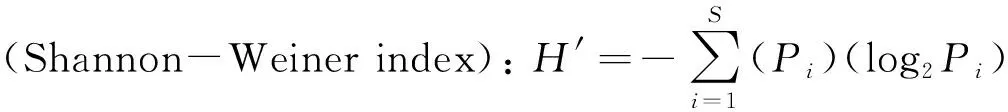

式中:Pi代表第i个物种数量占群落总个体数量的比例;S代表样地内的物种数;Hmax是最大香农-威纳指数。
1.4 土壤质量及草地综合质量评价

1.4.2草地质量综合评价 1)评价指标的确定。指标的确定通常根据4个原则来进行,即代表性、敏感性、实用性、独立性。也可使用主成分分析法,首先,选择主成分特征值大于1的主成分;然后,找出每个主成分中载荷最高的指标,并且选出载荷值在最高载荷90%以上的其余指标。若某个主成分的高载荷值指标只有一个,那么该指标被选取;若不止一个,将对其分别做相关性分析,相关系数小于0.7,各指标均被选取,若相关系数大于0.7,则选取相关系数之后最大的指标。

1.5 数据分析
对围栏内外的各项土壤理化性质数据、生物量及物种多样性数据分别进行正态性检验,若不符合则进行对数转换使其满足正态性,利用Bartlett Test检验其方差齐性。运用单因素方差(One way-ANOVA)分析,对比围栏与放牧处理下各指标造成的差异,所有均值检验数据以平均值±标准误(Mean±SE)的形式呈现。对土壤理化性质、生物量及物种多样性进行Pearson相关分析,所有检验均在α=0.05的置信水平下进行;利用主成分分析法对土壤质量进行评估,并筛选出关键性的综合评估指标。用SPSS 19.0(IBM SPSS Inc., Chicago, USA)进行统计分析,用Origin 8.5(OriginLab, Virginia, USA)和R语言软件包Corrplot作图。
2 结果与分析
2.1 围栏内外高山草地的土壤温度
图1显示随着试验样地的积雪逐渐融化殆尽,土壤温度随之逐渐提升,6-9月属于绝大部分植物的最佳生长时期。从日均温来看,围栏内外温度变化趋势相似,但围栏外温度波动幅度稍大于围栏内,7月下旬至8月下旬期间尤甚;日最高温数据显示,土壤温度在7月下旬至8月中旬这段时间内达到最大值,期间围栏外日最高温远高于围栏内,围栏内外日最高温分别为18.5和44.5 ℃;围栏内日最低温为6月初的4.5 ℃,围栏外的日最低温在8月初最低为-2 ℃;围栏内土壤每日温度最大波动幅度为7.5 ℃,而围栏外达43 ℃。在整个生长季内,围栏内土壤温度的日极值与日均温保持一致的变化趋势,而围栏外土壤温度日最大、最小值和日均值变化趋势并无一致性。
2.2 围栏内外的草地群落生物量和物种多样性
凋落物质量和地上生物量在围栏内外均有显著差异(P<0.05,表1),相对于围栏外的放牧处理,围栏内的凋落物质量和地上生物量分别增加了66.94%、35.77%。苔藓和维管束植物在围栏内的质量均大于围栏外,分别增加了56.25%和18.31%,但二者差异并不显著(P>0.05)。Shannon-Weiner多样性指数、Simpson多样性指数、Pielou均匀度指数及物种数在围栏内外均无显著差异(P>0.05)。
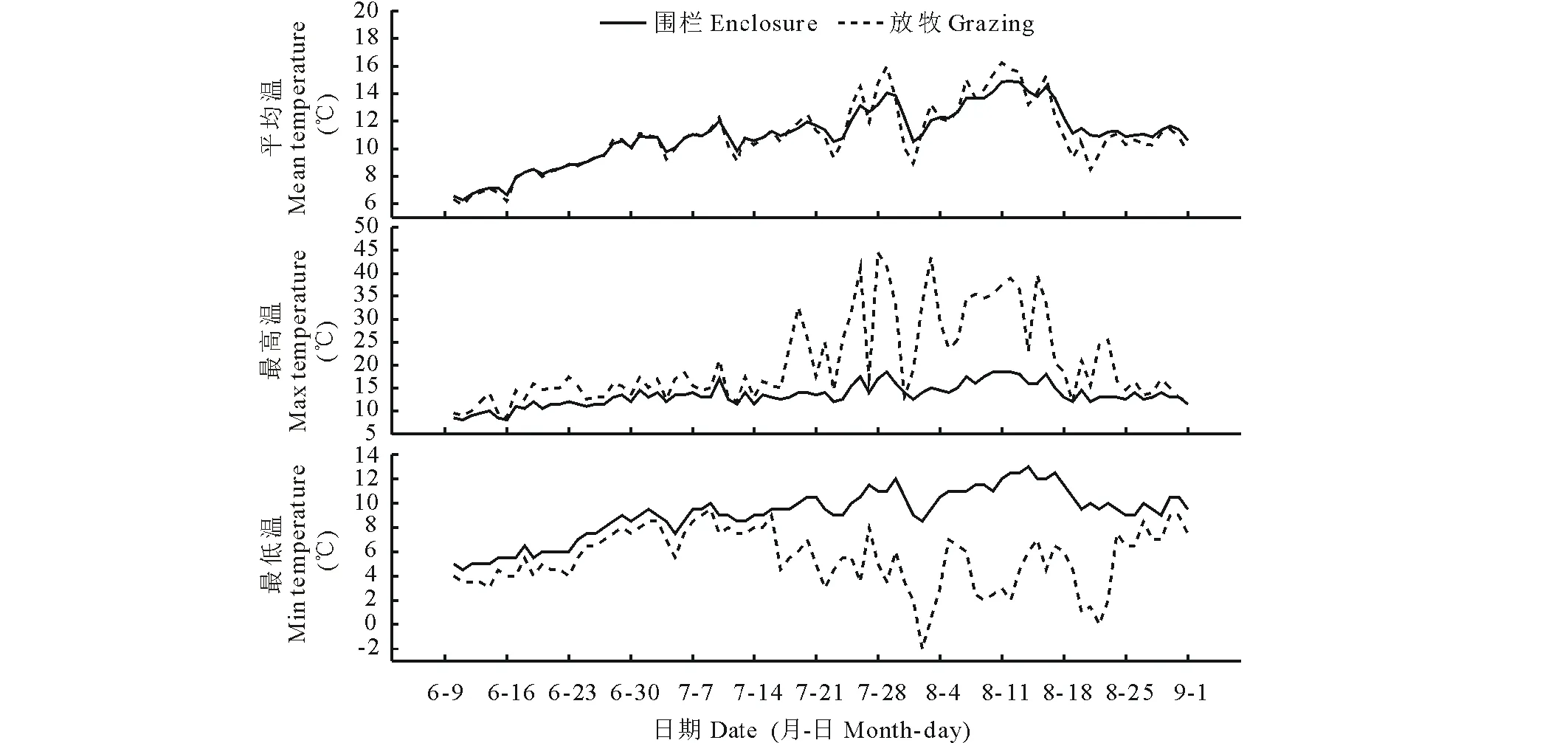
图1 生长季围栏内外土壤温度的变化Fig.1 The variation of soil temperature between enclosure and grazing during growing season
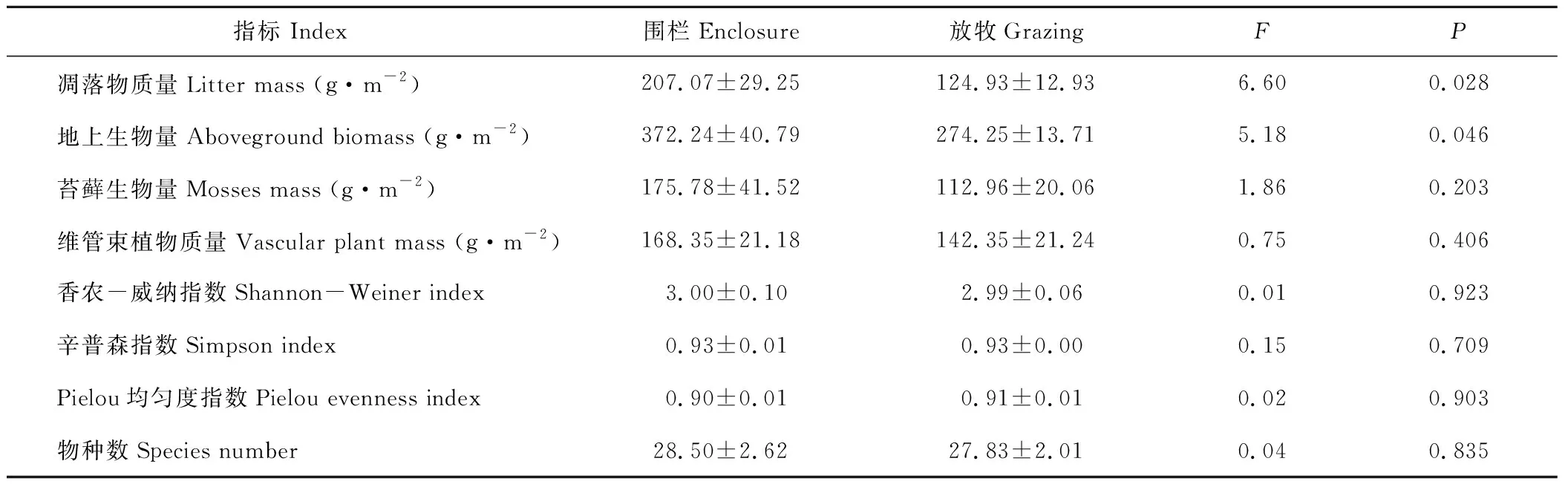
表1 围栏内外群落生物量及物种多样性比较Table 1 Comparison of community biomass and biodiversity between enclosure and grazing
2.3 围栏内外土壤理化指标的变化
土壤各项指标中(表2),仅硝态氮含量存在显著差异(P<0.05),围栏内的含量约为围栏外的2倍。相较于围栏外放牧处理,围栏内的土壤有机碳含量、全氮含量以及铵态氮含量和土壤含水量都有略微的提高,分别增加了9.19%、8.86%、7.25%、7.93%,但其差异均不显著(P>0.05)。土壤微生物生物量碳、氮含量及二者的比值在围栏内外均无显著差异(P>0.05),但围栏内存在下降趋势,分别减少了19.16%、7.22%、11.13%。此外,围栏内的容重降低了6.27%(P>0.05),其余土壤理化指标无明显变化。
2.4 高山草地土壤理化性质指标之间的相关性
对土壤理化性质各指标进行相关性分析(图2)。7年围封修复后围栏内外土壤理化性质相关性之间的差异明显。围栏内TK与ANC, TP与SWC呈显著正相关(P<0.05),TN与pH、MBN呈显著负相关(P<0.05),SBD与TP、SWC呈显著负相关(P<0.05);而围栏外土壤pH与TN, SBD与NNC, ANC与MBC/MBN均呈显著负相关(P<0.05),CEC与SOC、TN及TP均呈现显著正相关(P<0.05),而与pH呈负相关(P<0.05),MBC、MBN均分别与TN、TP呈正相关(P<0.05),同时MBN还与NNC及MBC呈现显著正相关(P<0.05)。

表2 围栏内外土壤理化性质比较Table 2 Comparison of soil physic-chemical properties at both sides of the fence
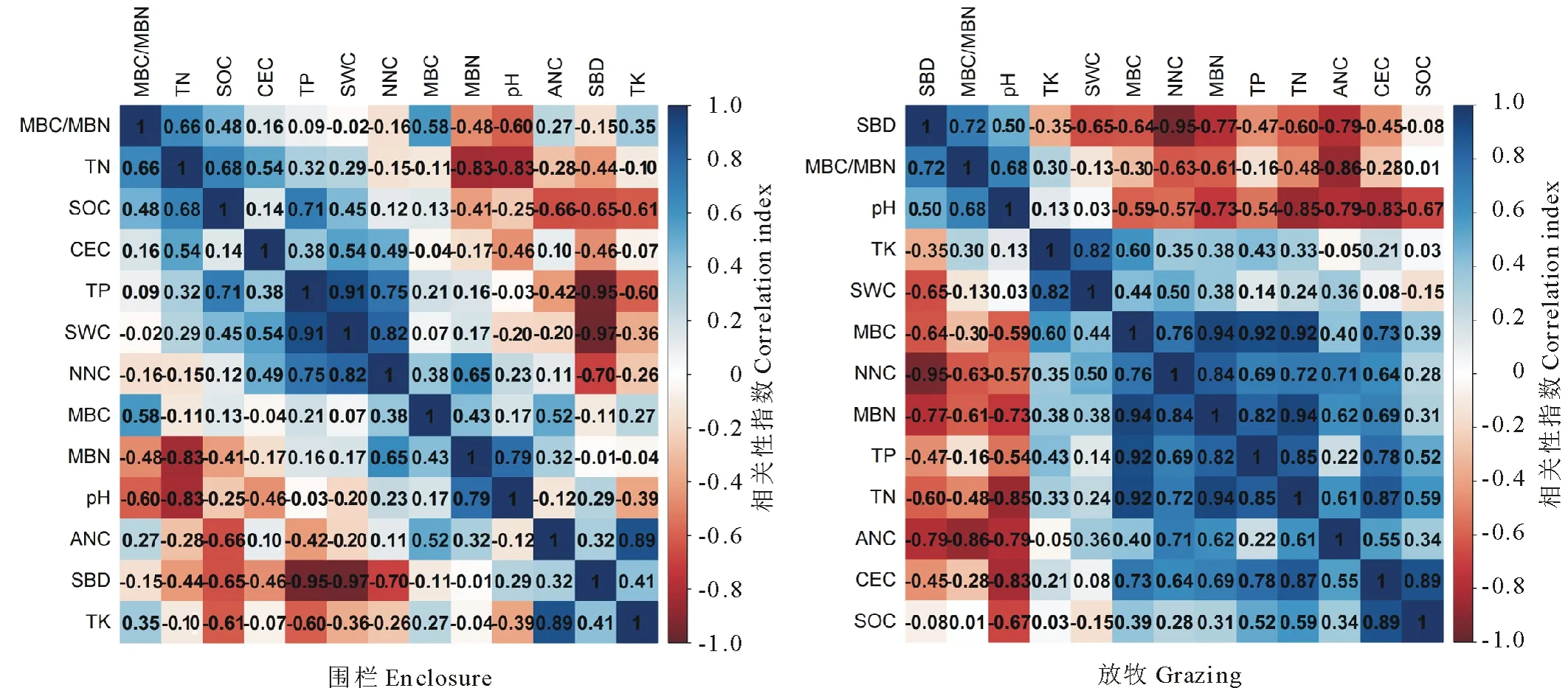
图2 土壤理化性质指标间的矩阵相关性Fig.2 Correlations of matrixes between different soil physic-chemical property indexes 蓝色表示正相关,颜色越深正相关性越强;红色表示负相关,颜色越深负相关性越强。Blue and red indicate positive correlation and negative correlation, respectively. The darker color presents the stronger correlation.
2.5 高山草地土壤质量评估
2.5.1土壤因子主成分分析 对上述13项土壤理化性质进行主成分分析,为了消除不同指标间量纲的不同以及数量级的差异,分析之前先将数据标准化(表3)。根据特征值>1,累计贡献率>85%的原则提取5个主成分,累计贡献率达88.86%,即这5个主成分可表达出所有土壤指标88.86%的信息。因此,采用主成分分析法是有效可靠的。一般认为某项指标的主成分载荷越高,那么它在该主成分中所占权重越大。由此可见,第1主成分以SBD、TN、NNC为主要影响因子;第2主成分以TK、MBC、SOC为主要影响因子;第3主成分以MBC/MBN为主要影响因子;第4主成分则以ANC为主要影响因子;第5主成分以SWC为主要影响因子。

表3 土壤因子的主成分值和特征向量Table 3 The principal component and eigenvector of soil factors
注:EV, 特征值; CR, 贡献率; CCR, 累积贡献率。
Note:EV, Eigenvalue; CR, Contribution rate; CCR, Cumulative contribution rate.
2.5.2特征向量和各主成分方程 主成分的各特征值见表3。根据特征值和主成分的载荷值得到每个主成分的特征向量(方法见1.4.1)。再依据特征向量建立主成分方程。5个主成分的方程为:F1=-0.298pH+0.326CEC+0.238SOC+0.357TN+0.331TP+0.081TK+0.335NNC+0.193ANC+0.242MBC+0.286MBN-0.083MBC/MBN+0.272SWC-0.365SBD,F2、F3、F4、F5以此类推。
2.5.3主成分得分、综合得分 各主成分得分及土壤综合得分见表4,从表中可以看出8号样的综合得分最高(1.95),9号样最低(-1.26)。围栏内的土壤样本平均综合得分高于围栏外且较为稳定,其平均得分为0.1,围栏外平均得分为-0.02,经方差分析发现围栏内外得分差异不显著(P>0.05)。
2.6 综合评估围栏措施对高山草地质量的影响
草地质量分为地上部分与地下部分,地上部分主要是指植被质量,地下部分主要指土壤质量。根据指标选取的4个原则(代表性、敏感性、实用性、独立性),综合考虑选取的地上部分指标为凋落物质量、苔藓生物量及Simpson多样性指数。地下指标依据主成分分析结果选择(见1.4.2),第1主成分高因子载荷指标有:TN、TP、NNC和SBD;第2主成分高因子载荷指标有:TK; 第3主成分高因子载荷指标有:MBC/MBN;第4主成分高因子载荷指标有:TP、ANC;第5主成分高因子载荷指标有:SWC。第4主成分高因子载荷指标有2个,二者相关系数低于0.7,因此均被选取,第1主成分的NNC与SBD相关系数大于0.7,且SBD的相关系数总和较高应被选取,但考虑到选取指标的敏感性原则,因此NNC也被选取。以上11个指标被选取进行草地质量综合评价,各指标的权重确定根据相关系数法(见1.4.2),由围栏内外所有样本数据计算得到。评价结果见表5。

表4 样本的主成分得分及综合得分Table 4 The principal component value and comprehensive value of principal component of samples
注:1~6号为围栏内土壤样品,7~12号为围栏外土壤样品,Fi(i=1,2,3,…)为各主成分得分,FZ为综合主成分得分。
Note: No.1 to 6 are samples from enclosure, No.7 to 12 are samples from grazing,Fi(i=1,2,3,…)are the principal component value respectively,FZis comprehensive value of principal component.

表5 高山草地质量综合评价Table 5 The comprehensive assessment of alpine meadow quality
3 讨论
实施围栏管理是修复和防止草地退化的重要手段之一,在全球范围内有着广泛的应用[23,34]。与大多数研究结果类似,本研究发现退化高山草地围栏7年后,围栏内土壤性质得到一定程度的改善[24,26]。首先,土壤温度方面,生长季盛期的围栏内外土壤日均温波动均较大,主要是强烈的太阳辐射以及超过70%的年降水均发生在此阶段。长期放牧导致围栏外其日最高、最低温波动幅度较围栏内大,主要是因为在围栏内的植被长势较好,下垫面对温度变化起到一定的缓冲作用。而围栏外存在一些非生物因素(主要为放牧)的干扰,导致植被的丰富度及均匀度降低,地表土壤直接接受了更多的太阳辐射,同时围栏外土壤含水量较低且土壤结构紧实,导致其反照率及土壤热传导率较高,夜间热量散失较快[35-36]。另外,7年的围封使得围栏内土壤腐殖层的厚度增加,其对温度的隔绝性也较高,因此围栏内土壤温度日极值变化较小。对土壤理化指标间的相关分析发现,围栏的修复过程给土壤介质各理化指标及其相关性影响很大。首先,土壤物理性质方面,围栏的设置有效地减少了牲畜对表层土壤的践踏,使土壤在无外界干扰的情况下逐渐自我修复。土壤孔隙度增加并疏松化,围栏内土壤含水量及容重指标得到优化。同时,围栏也有效地防止了牲畜对植被的破坏,植物群落的生长有利于土壤团聚体的形成,促进土壤物理性质的改善[37];土壤化学性质方面,围栏内的土壤有机碳、全氮及铵态氮的含量均有小幅度增加趋势,主要是因为土壤及植被得到了围栏的有效保护,凋落物质量显著提高,可能促使有机质分解作用增加,促进了微生境内的生物地化循环[38]。7年的围栏修复后,围栏内外的土壤硝态氮含量存在显著的差异。可能有3个原因:1)硝态氮主要来源于铵态氮的硝化作用,围栏内的铵态氮含量相较于持续放牧处理提高了7.25%,硝化作用底物积累更有利于硝化产物的增加;2)相关分析表明土壤硝态氮含量与土壤含水量呈正相关关系,与土壤容重呈负相关关系,围栏内的土壤含水量提高了7.93%(表2),也许更利于硝态氮离子形态的积累;3)围栏内土壤容重得到改善,土壤孔隙度提高、空气含量增加促进硝化作用,亦使得硝态氮含量升高。微生物生物量碳、氮含量也许与放牧区牲畜的活动有关,其排泄物对土壤表层的营养物质会造成一定影响。从总体来看,围栏对土壤理化性质的改善具有积极作用,但也有研究发现围栏也存在负影响机制,例如由于围封草地凋落物过多导致其分解速率下降,不利于物质循环过程[13]。因此,在实际应用中要充分考虑围栏对于退化草地生态系统修复作用的局限性以及时空特异性。
退化草地围封7年后其植被得到修复,其中凋落物质量和地上生物量有显著的提高,苔藓和维管束植物生物量也趋于增加,这主要是由于围栏有效地防止了牲畜采食,同时一定程度上土壤理化性质的改善与植物的生长有着良性互馈。此外,围栏内外的物种多样性及物种数变化并不显著,这与前人的一些研究结果类似[11,39],可能与围封时间、土壤质量、群落结构以及特殊的地理位置等相关。以往的研究结果表明,围栏对物种多样性造成的变化趋势不尽相同。一些研究者认为,围栏的保护机制让草地植物规避了人为干扰因素,物种多样性得以恢复,但恢复效果与围栏时间之间并非简单的线性关系[40-41];而另一些研究者认为,围栏恢复具有强竞争力物种的生长压力后,资源限制区域内竞争力较弱的物种开始消失,从而导致物种多样性逐步降低[39,42-43]。综上,对于群落结构相对简单的高山草地系统来说,维持稳定的草地生物量产量、保护物种生物多样性对其可持续发展与利用具有重要意义。
高山草地土壤质量状况数值化的综合评价,能更直观地以质量综合得分为依据来判断草地保护及资源使用的情况[44]。本研究土壤质量综合得分最高(1.95)和最低(-1.26)的土壤样品均来自放牧区(均值为-0.02),其平均值略低于围栏内(0.1)。尽管二者的方差分析结果表明二者无显著差异,但是放牧区综合得分比围栏内的波动大,即表明7年内围栏较稳定地改善了土壤质量,而围栏外的草地土壤质量呈现斑块化的分布,这可能是由牲畜选择性采食造成的。围栏内草地综合质量亦有小幅度的提高(相对得分0.209),草地综合质量中土壤质量占了较大的比重(78%),其中土壤含水量、容重及硝态氮含量的权重较高,直接与土壤的水肥气热相关。因此,草地保护过程中既要注意土壤质量的修复情况,同时也要考虑地表植被的恢复状况。此外,可视其退化程度辅以开展松土或提高相应肥力的措施,并且有必要开展定期的草地恢复监测工作,并可进行围栏修复的时序性研究便于对其时效性做出正确判断。草地生态系统弹力恢复后可进行人工刈割及低强度放牧活动,在放牧策略上也可选择季节性放牧或轮牧方式以避免围栏带来的负面影响。
4 结论
围栏是管理草地的一种人为措施,其目的在于科学地平衡生产与利用之间的关系。本研究发现严重退化的高山草地在经过7年的围栏封育后,围栏内大部分土壤理化性质相较于持续放牧处理有所改善,以硝态氮含量变化最为显著。地上部分凋落物质量和地上生物量显著增加,物种多样性方面尚无显著性变化。草地综合质量评价表明,土壤质量有小幅度的提高,土壤含水量、土壤容重以及氮、磷元素是草地恢复中的重要指标,土壤质量的优劣对于草地质量的提高非常重要。采取草地质量综合评估法探讨高山草地的修复效果,旨在为草地修复情况的判定提供一种科学有效的手段,在一定程度上对高山草地的修复工作具有科学的指导意义,而在其他区域或更大时空尺度的适用性尚待进一步探讨。
