圈养马麝(Moschus sifanicus)个性特征及与麝香分泌的关系
2018-12-19孙太福孟秀祥
周 杨,孙太福,黎 勇,周 密,孟秀祥,,*
1 中国人民大学环境学院,北京 100872 2 兰州逢春麝业科技有限公司,榆中 730116 3 山西农业大学动物科技学院,太谷 030801
个性(Personality)通常是指不同时空下同种生物个体间行为特征的稳定差异[1-2]。20世纪80年代左右,行为生态学家开始关注动物个体间个性差异问题[3],针对无脊椎动物[4]、鱼类[5]、鸟类[6]、哺乳动物[7]等动物类群均进行了广泛研究。动物的个性在年龄、性别间存在差异,并具有一定的遗传性[8]。不同于人类个性研究,动物个性研究一般通过镜子实验、旷场试验、新异物实验等方式,获取行为实验数据,进行个性维度划分,如侵犯性、胆怯性、社会性、活跃性、领域性、探索性等[9],个性特征的不同程度以及在不同情境中的变化与动物觅食、繁殖、扩散、反天敌等生活史对策之间紧密联系[10]。个性特征研究已经逐渐成为动物行为生态领域的热点,近5年相关成果不断增加[11]。众多研究多集中于模式动物或小型实验室动物[12],如Cote等[13]通过对食蚊鱼的研究探讨了在捕食压力下的动物个性扩散行为,哺乳类动物多集中于啮齿类小型动物,如程琪等[14]对高原鼠兔个性特征的地理变异进行了分析。受限于样本数量和实验控制条件,有蹄类动物中鲜见动物个性的量化研究,少量相关研究一般通过行为观察取样来实现,如Jennings等[15]研究了黇鹿个性及打斗行为与繁殖成功的关系,Strong等[16]对雄性驯鹿个性与交配行为的关系进行了分析,而在类似马麝的小型有蹄类动物中目前尚无个性方面的量化研究。
马麝(Moschussifanicus)为森林小型哺乳类,为我国一级保护动物,成年雄麝所分泌麝香被广泛用于亚洲传统医药和香料行业,过度利用及生境恶化造成马麝种群数量急剧下降以至濒危[17-18],当前主要通过圈养方式对其进行迁地保育[19]。马麝行为学在个体活动节律、刻板行为、行为多样性及行为格局等多方面均有全面深入的研究[19-20],但因种群数量有限,且较长一段时间里马麝的圈养并未达到一定规模,饲养管理模式不统一,不具备良好的行为取样和实验控制条件。不同于实验性质的动物,小型哺乳动物个性研究无法进行严格的控制实验,所以通常使用个性量表评分法进行,如对工作犬的个性特征研究等[21],而马麝作为圈养动物,动物饲养管理复杂,较难实施量表评分标准化测试,且带有一定主观性。
20世纪90年代开始,本研究所在麝场开展圈养马麝实践,经过多年发展,形成了具有规模的标准化管理模式,存圈数达500头,成为濒危珍稀保护动物迁地保育的优秀范例。基于此,本研究所涉及马麝个体样本量充足,且具有良好的取样和实验条件,使个性研究成为可能。通过对动物进行独立的行为取样,规避量表评分法缺点,同时能得到类似实验室动物个性研究的行为学标准化数据[4],是类似的小型有蹄类个性研究的一个可行方案。马麝个性特征与生活史事件相关联,并在年龄和性别间存在一定差异,通过对马麝个性的量化,是分析个性之间相互关系及不同情境季节间的个性特征变化的前提,进而探讨不同个性特征对雄麝泌香量的效应。对马麝个性特征的量化研究,不仅对马麝个性行为学理论创新有参考价值,也为麝香产量预测和优质品种选育配种提供了一种可行方案,对麝香及麝类资源的发展具有重要实践意义。
1 材料与方法
1.1 研究地点及实验动物
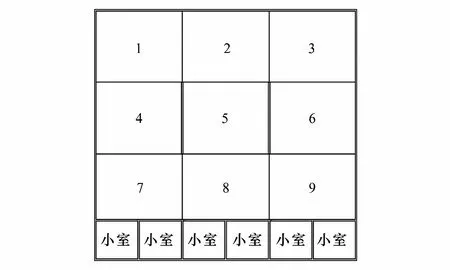
图1 麝场圈舍结构及位点划分Fig.1 A plan of the enclosure
本研究于甘肃省兴隆山自然保护区马麝繁育试验场(以下称麝场)进行。麝场所在区域地理、气候特征及动物繁育管理情况详见文献报道[22]。本研究涉及麝场23个标准独立圈舍,内部结构设施一致,每个圈舍包含若干小室(cell)和1个面积为10 m×10 m的外部活动场区域,活动场基底均为泥质裸地,中央区域建有遮阳棚,供饲喂及动物休憩使用,将外部活动区域按九宫格形式划分为9个位点,其中5号位区域为中央凉棚区域[23](图1)。
本研究共涉及81头圈养马麝(雄性28头,雌性53头),圈舍内包含一雄多雌,个体情况如表1所示(表1)。将个体按幼年个体(1.5—2.5岁)、成体(3.5—6.5岁)、老年个体(7.5岁及以上)进行年龄组区分。兴隆山麝场始建于20世纪90年代初,具有完备的雄麝泌香相关数据库,得到目标雄麝年均泌香重量(g)(2008—2017)作为雄麝泌香水平的标准。
1.2 研究方法
行为是指动物在一定环境条件下,为完成摄食、排遗、体温调节、生存繁殖及满足个体其他生理需要而以一定姿势完成的一系列动作,结合行为谱研究文献[24-25],建立本研究马麝行为谱,对个体进行行为取样,并从中提取行为进行个性特征的区分。麝场马麝个体均安装唯一塑料耳号,用以快速个体识别,利用肉眼和10×50°望远镜协助行为观察。兴隆山马麝交配行为始于11月中旬,本研究取样时间为2016年11—12月和2017年7—8月,分别为交配季节与非交配季节[26-27],取样时间段为07:00—12:00和14:00—20:00,所有取样均由一人完成[24]。
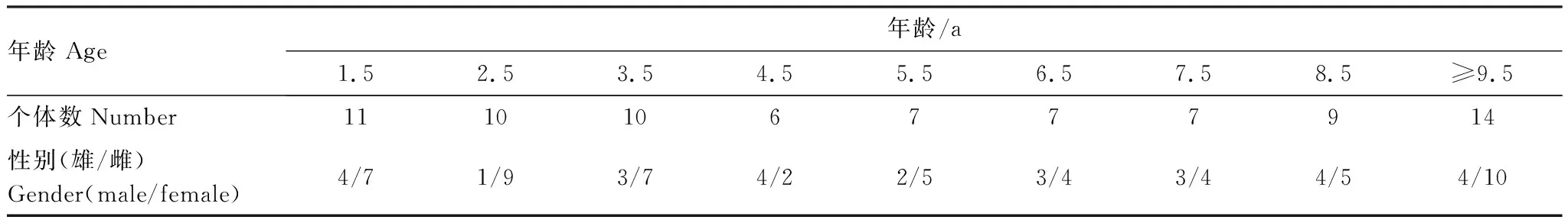
表1 目标马麝全部个体数统计
参照动物行为取样研究文献[28],对马麝个体进行焦点行为取样(Focal sampling)和扫描行为取样(Scan sampling)。焦点行为取样记录个体行为类别、发生时刻、发生位点,每次取样持续时间为10 min,若个体脱离观察者视线,则该行为样本不参与统计分析。对全部圈舍进行扫描行为取样,设置扫描时间间隔为1 h,避免扫描取样过程中个体产生惊扰而未恢复行为常态,若扫描过程中个体脱离观察者视线,记为空白值,不参与统计分析[29]。
结合行为谱和行为取样结果,对行为样本进行整理。将扫描取样样本中非静息行为(除“静卧”、“静卧反刍”、“卧睡”之外的行为)发生比例作为马麝活跃性的量化指标;将焦点取样过程中刻板行为类行为(食异物、搭蹄观望、往返走、狂奔、刻板舔舐、跳墙、立台)和环境探究行为类行为(嗅墙、嗅地、嗅蹭尾处、嗅食物、嗅便所、嗅卧迹)的发生强度(单位时间内发生频次)进行标准化,衡量马麝刻板性水平及环境探索性水平;参照动物行为冗余性文献[19,30],计算行为冗余性,衡量马麝行为冗余的程度;圈舍内1、3、7、9位点为四角位置(图1),是圈舍内个体占据的热点领域,通过焦点取样中以上位点的占据时间来衡量个体领域性水平。
活跃性指数=非静息行为发生频次/扫描取样次数×100%
刻板性指数=刻板行为发生频次/600 s×100%
环境探索性指数=环境探索行为发生频次/600 s×100%
行为冗余性指数=1-行为多样性指数r[19]
领域性指数=圈舍热点区域占据时间/600 s×100%
1.3 数据分析
采用Kolmogorov-Smirnov Test对个体个性特征及雄麝泌香量进行正态性检验,如数据呈正态分布,则采用相应的参数检验方法(Parametric test)检验各目标因子效应。采用Pearson相关(双侧)分析各个性特征之间及与泌香量之间关系;采用One-way ANOVA分别检验交配季节及非交配季节内个性特征在年龄组及性别之间的差异;对交配季节及非交配季节之间个性特征进行配对T检验(Paired ‘matched’ t-test)及Pearson相关(双侧)分析。本文设定差异显著性标准为α=0.05,所有数据分析通过SPSS 19.0进行。
2 结果
2.1 性别及年龄组间个性特征差异
交配季节与非交配季节里,马麝活跃性、刻板性、环境探索性、行为冗余性、领域性、泌香量指数均呈正态分布(表2)。
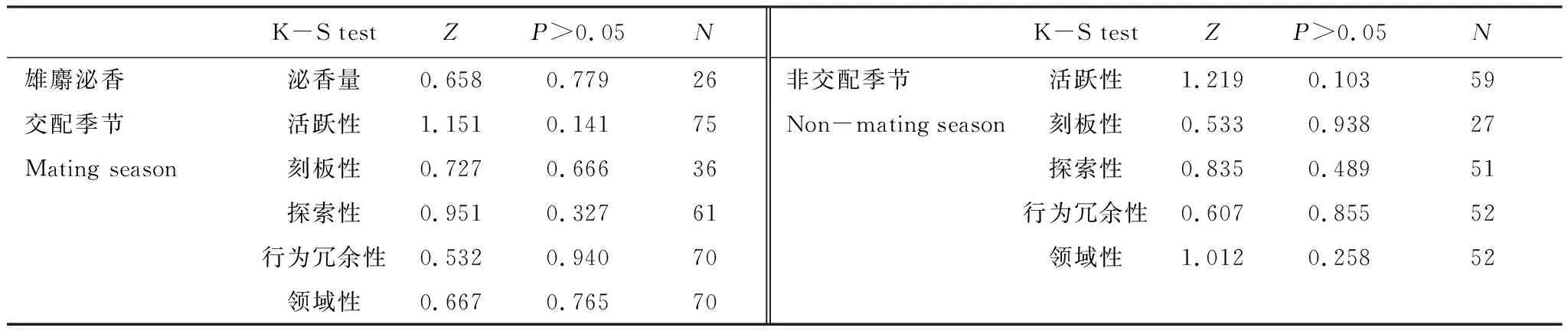
表2 正态性检验
交配季节里,活跃性随年龄增加有显著降低的趋势(ANOVA,F2,74=4.393,P=0.016<0.05),刻板性(P=0.386>0.05)、探索性(P=0.516>0.05)、行为冗余性(P=0.164>0.05)及领域性(P=0.145>0.05)均无显著差异;雌雄之间活跃性(P=0.434>0.05)、刻板性(P=0.339>0.05)、探索性(P=0.186>0.05)、行为冗余性(P=0.664>0.05)及领域性(P=0.755>0.05)同样无显著差异(图2)。
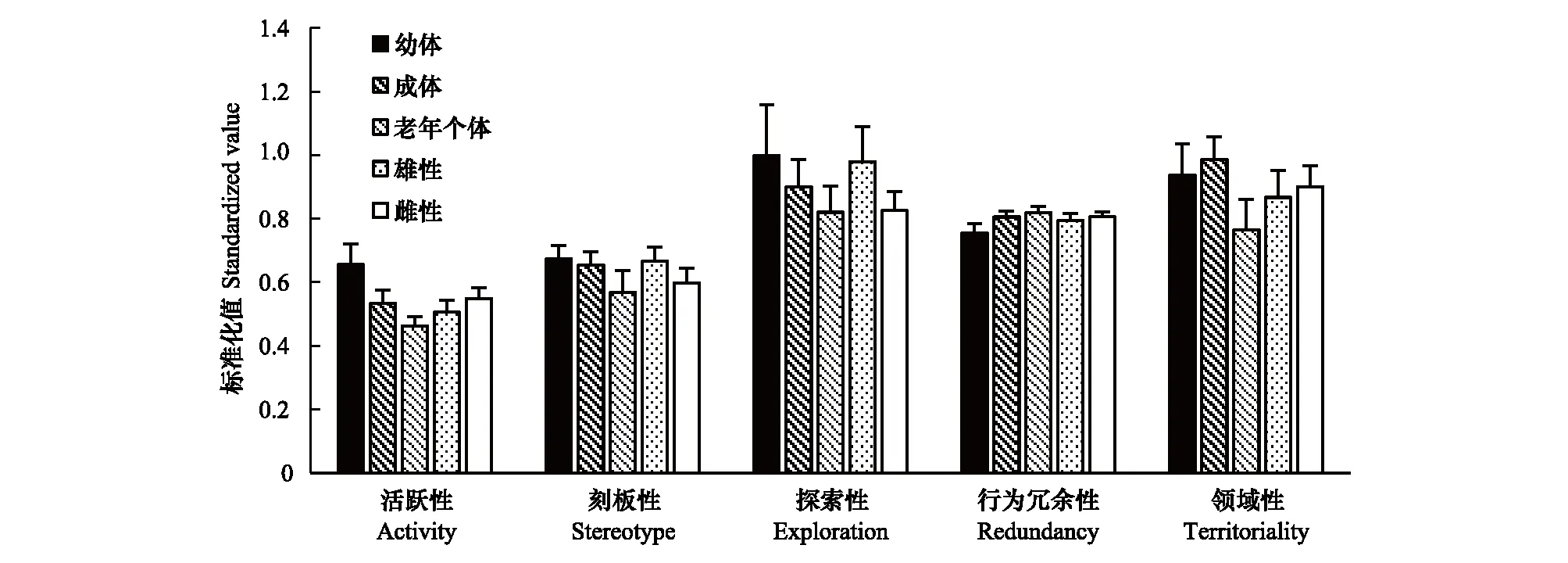
图2 交配季节个性特征差异Fig.2 Difference of personality traits in mating season
非交配季节里,活跃性(ANOVA,F2,58=4.476,P=0.016<0.05)与领域性(ANOVA,F2,51=3.881,P=0.027<0.05)随年龄增加均有明显降低趋势;行为冗余性随年龄增加具有升高的趋势,差异接近显著(ANOVA,F2,51=2.874,P=0.066>0.05);各年龄组探索性(P=0.74>0.05)、刻板性(P=0.243>0.05)差异均不显著。雌性活跃性显著高于雄性(ANOVA,F1,58=8.527,P=0.005<0.05),但刻板性(P=0.328>0.05)、探索性(P=0.329>0.05)、行为冗余性(P=0.716>0.05)、领域性(P=0.982>0.05)差异不显著(图3)。
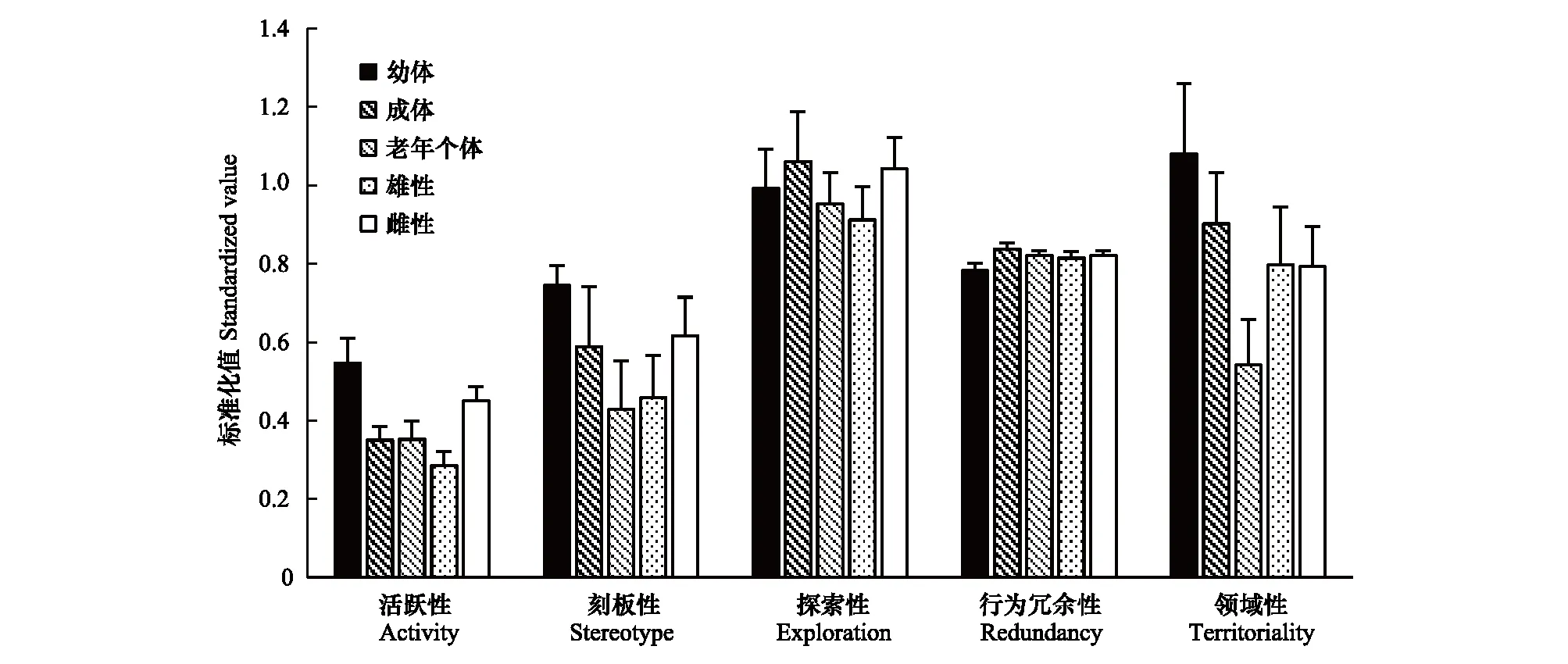
图3 非交配季节个性特征差异Fig.3 Difference of personality traits in non-mating season
2.2 交配季节与非交配季节间个性特征比较
雄性个体,交配季节活跃性(0.493±0.05)极显著高于非交配季节(0.287±0.04)(P=0.000<0.01);交配季节刻板性(0.647±0.05)高于非交配季节(0.542±0.21),差异不显著(P=0.720>0.05);交配季节探索性(0.883±0.15)高于非交配季节(0.816±0.09),差异不显著(P=0.688>0.05);交配季节行为冗余性(0.814±0.03)与非交配季节(0.815±0.02)差异不显著(P=0.962>0.05);交配季节领域性(0.814±0.12)高于非交配季节(0.798±0.15),差异不显著(P=0.897>0.05)(图4)。
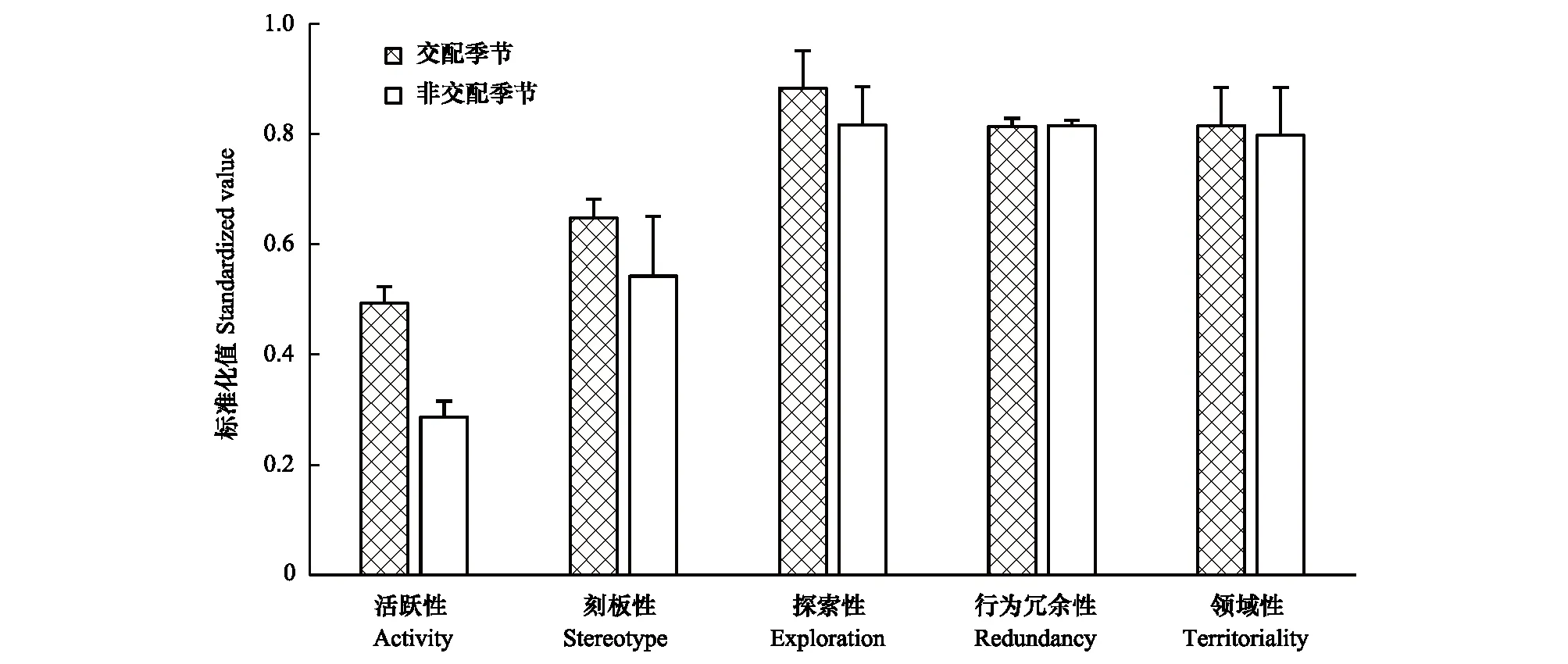
图4 交配季节与非交配季节雄性个性特征比较Fig.4 Comparison of personality traits of males between mating season and non-mating season
雌性个体,交配季节活跃性(0.516±0.04)高于非交配季节(0.424±0.04),接近显著(P=0.051>0.05);交配季节刻板性(0.675±0.04)略低于非交配季节(0.707±0.13),差异不显著(P=0.843>0.05);交配季节探索性(0.812±0.07)显著低于非交配季节(1.094±0.09)(P=0.026<0.05);交配季节行为冗余性(0.793±0.02)低于非交配季节(0.820±0.01),差异不显著(P=0.196>0.05);交配季节领域性(0.864±0.09)高于非交配季节领域性(0.750±0.11),差异不显著(P=0.314>0.05)(图5)。
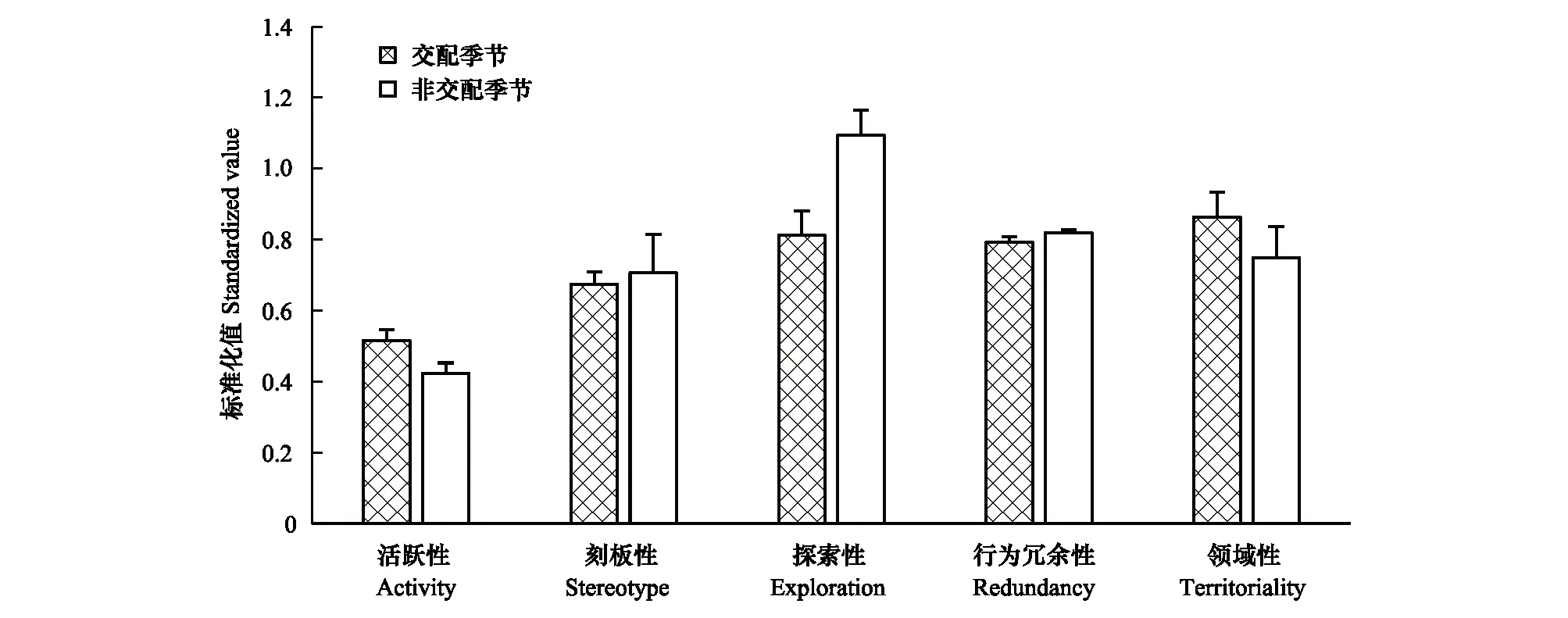
图5 交配季节与非交配季节雌性个性特征比较Fig.5 Comparison of personality traits of females between mating season and non-mating season
雄性交配季节活跃性与非交配季节呈显著正相关关系(r=0.504,P=0.028<0.05,N=19),但在雌性中并不显著(P=0.115>0.05);交配季节领域性与非交配季节呈显著正相关关系(雄性:r=0.547,P=0.028<0.05,N=16;雌性:r=0.372,P=0.047<0.05,N=29);交配季节刻板性与非交配季节呈显著负相关关系(雄性:r=-0.986,P=0.014<0.05,N=4;雌性:r=-0.599,P=0.039<0.05,N=12);探索性(雄性:P=0.574>0.05,N=14;雌性:P=0.701>0.05,N=27)、行为冗余性(雄性:P=0.223>0.05,N=16;雌性:P=0.77>0.05,N=29)在两季节间无显著相关关系。
2.3 马麝个性特征之间相关性及与泌香量的关系
个体活跃性与行为冗余性呈显著负相关关系(交配季节:r=-0.239,P=0.047<0.05,N=70;非交配季节:r=-0.325,P=0.021<0.05,N=50);非交配季节里刻板性与探索性存在显著正相关关系(r=0.675,P=0.000<0.01,N=26)。
雄麝泌香量与活跃性存在较强的正相关性(交配季节:r=0.518,P=0.008<0.05,N=25;非交配季节:r=0.397,P=0.092>0.05,N=19),同时与交配季节领域性也存在显著的正相关关系(r=0.406,P=0.044<0.05,N=25)(图6;图7)。
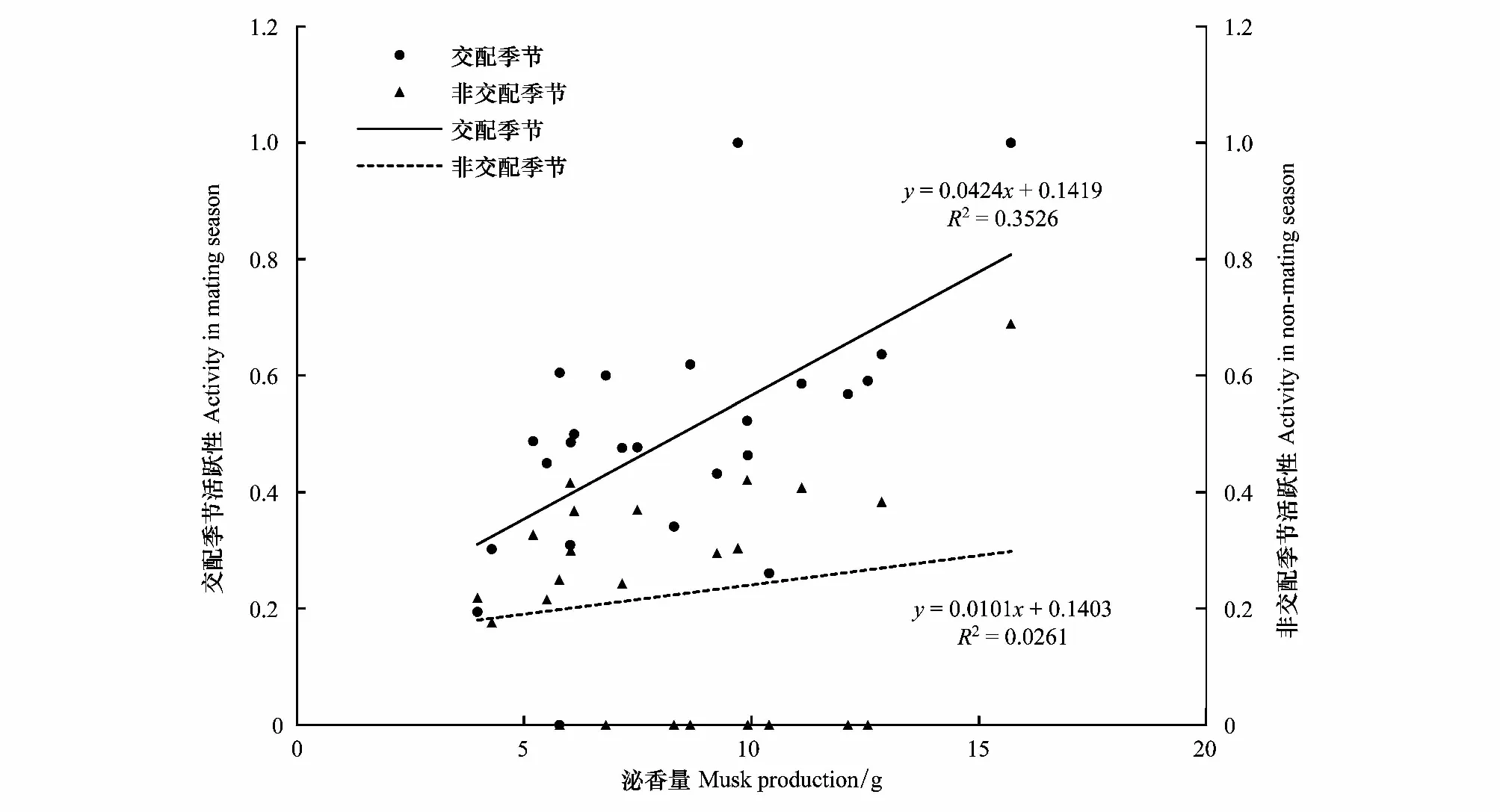
图6 泌香量与交配季节及非交配季节活跃性的关系Fig.6 Relationship between musk production and activity during mating season and non-mating season
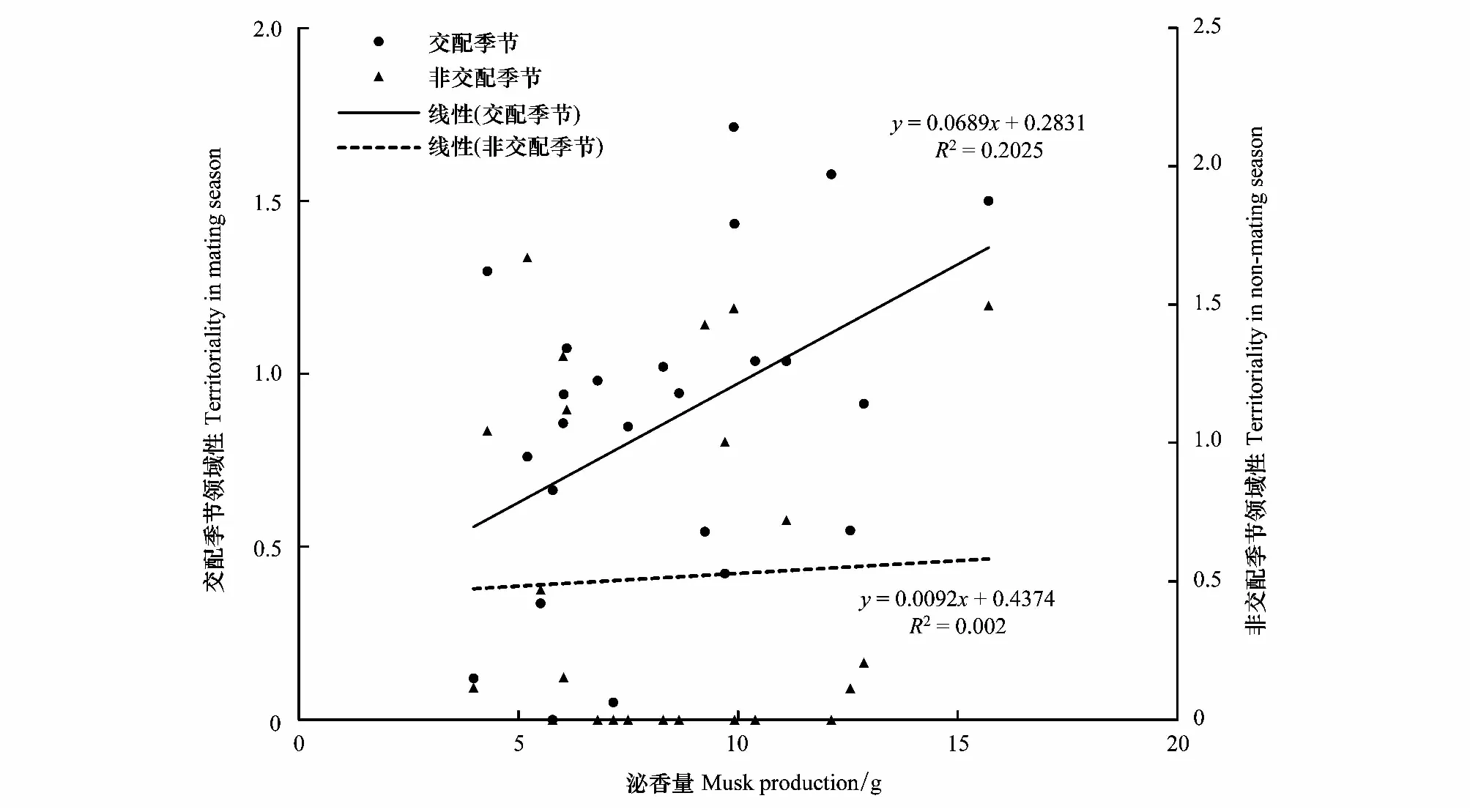
图7 泌香量与交配季节及非交配季节领域性的关系Fig.7 Relationship between musk production and territoriality during mating season and non-mating season
3 讨论
本研究所在麝场马麝年龄基本能代表其被圈养时间[31]。圈养条件下,马麝圈养时间越久,健康状态随之降低,单一的圈养环境和刻板行为的发育降低了个体行为多样性,行为冗余性上升[19]。Conrad等[4]认为活跃性是动物个体在觅食能量消耗与觅食收益之间的权衡,更高的的活跃性可能与更高的代谢率相关联。交配季节与非交配季节中,活跃性随年龄增加均具有降低的趋势,说明年龄(圈养时间)加剧了个体的行为冗余性,对动物个体健康度产生一定影响,从而活跃性也相应降低,这也符合马麝个体的一般生长规律,年老个体代谢率一般趋于下降,本研究同样得到了随着个体年龄增加,非交配季节行为冗余性明显升高,佐证了上述结论。交配季节与非交配季节刻板性和探索性同样随着年龄增加具有降低的趋势,证明圈养时间不仅对刻板行为的发育产生效应,也降低了个体环境探索性行为的发生,推测更大的圈养面积和更高的圈养环境丰富度有益于圈养动物福利,促进自然行为发育[19]。本研究中雌性活跃性在非交配季节显著高于雄性,主要由于雌麝非交配季节处于产后恢复期和哺乳期,警戒性增强,行为更多表现为站立凝视和摄食,而雄麝更多通过卧息行为来减少消耗,为繁殖季节储能[29]。与此同时,雄麝在非交配季节正处于泌香期,泌香过程中出现食量减少,活动量下降等生理现象,相较于同时段雌性个体,雄性具有更低的活跃程度[32]。
非交配季节期间个体领域性随年龄增加具有明显降低的趋势,一方面是因为圈舍内特殊位点(四角)相应具有更高的安全性,能警戒更大的空间范围,同时接近隐蔽小室或食物投放点,是较为优势的区域位点,老年个体领域性水平较低可能与其健康度和等级序位相关联;另一方面,与赤腹松鼠在繁殖季节具有较强巢域不同[33],马麝繁殖交配季节并未体现差异化的领域性,这是由于圈养条件的限制使雌雄个体交配过程更多的需要通过动态位移来完成,而在非交配季节里,初次繁殖个体对特殊位点的占据更加强烈,圈舍四角为警戒观察、奔跑、跳跃等邀配行为所发生的重点区域。
动物个性特征伴随整个生活史过程,可以看作是一个不稳定的生活史特征,并在相应的季节情境间产生变化[34]。马麝个体交配季节活跃性明显高于非交配季节,交配季节个体处于发情期,更多的展现相互打斗或邀配行为,而在非交配季节里,雌麝个体处于产仔期,相较于交配季节期间进行大量摄食活动来储备能量,非交配季节活跃度相对有所下降,雄麝更多表现静卧等静息行为,在泌香过程中还会因生理原因造成活跃度下降,这与孟秀祥等[32]关于不同来源的雄性马麝行为差异研究相符,雄性个体交配季节静卧行为发生频次显著低于非交配季节。本研究中同样得到雌性马麝交配季节环境探索性显著低于非交配季节,符合兴隆山雌性马麝更多表达环境探究和尾阴探究等行为的结论,非交配季节更高的探索性是雌性收集环境信息和其他个体性信息的一种表现[29]。个性特征的变化是个体在不同情境下对环境因子的响应,虽然本研究中探索性、刻板性、行为冗余性并未探察到显著差异,但从经验角度的判定,圈养马麝在不同季节里的生活史过程对个性特征的影响是明显存在的。
Sih等[11]认为动物个体行为差异在不同情境间存在一致性,动物行为在跨情境间的组合进化称之为行为综合征现象。本研究中雄麝活跃性在不同季节间存在显著正相关关系,说明活跃性水平高低作为一对行为轴具有跨情境一致性[11]。与活跃性类似,领域性在两季节间的显著正相关关系说明了领域性和活跃性可能均为个体健康度和圈养环境中等级序位的一种体现,更加活跃的个体或占据优势区域的个体在交配季节里的表现更加良好,同时能延续到非交配季节中,这类似Johnson[35]对蜘蛛类群胆量个性的研究,认为个体在不同情境间调整自身胆量水平,且跨情境保持一致性,从而能进行情境间胆量水平的预测。与活跃性和领域性不同,马麝高刻板性对动物繁殖成效和生产力均会产生负面效应,定时限饲、圈养空间限制等是导致刻板行为发育和表达的重要因素[31]。本研究表明交配季节的刻板性水平与非交配季节呈负相关关系,而兴隆山马麝性活跃度更高的个体在交配季节展现较低水平的刻板行为表达[31],推测马麝刻板性特征同样具有行为的可塑性和表达弹性,性活跃度较高的个体在交配季节刻板性表达低,在非交配季节情境里,受饲养条件和圈舍环境限制的反馈强度更大,表现出更高水平的刻板性。
动物个性特征与适合度存在紧密联系[16],马麝泌香量能较好代表个体内分泌机能和生理功能的状态[36]。兴隆山马麝泌香量与交配季节活跃性和领域性正相关,这与个体的季节性行为模式相关联。交配季节,雄麝个体卧息行为较少,更多表达冲突性行为,运动及蹭尾等信息标记行为更多表达[29]。交配季节的活跃性和领域性一方面代表了个体本身健康度良好,等级序位相应更高,另一方面,更高的活跃性和领域性也带来了更多觅食资源回报,在交配季节里更充足的能量储备使非交配季节相应具有更多能量用以麝香分泌。马麝个体活跃性与行为冗余性的负相关关系佐证了个性特征与泌香量的关系,更高活跃度的个体行为冗余性更低,可能更加适应圈禁环境,相应的泌香量更高,健康状态更佳[19]。因此,交配季节里马麝个体的活跃性和领域性能作为雄麝泌香量的一种预测手段,能较为可靠的对雄麝种质质量进行评判,进一步的定量分析能达到通过个性特征值对雄麝泌香量进行较为准确的计算。马麝个性特征的研究是马麝行为学的补充,是麝香产量预测的一个可行方案,对雄麝选种配对和提高麝香产量具有实践意义。
致谢:感谢甘肃兴隆山麝场工作人员对本研究的支持。
