舟山海岛地区新重组GII.P16-GII.2型诺如病毒基因特征分析
2018-12-13,,1,,,,,
,,1,,, , ,
诺如病毒是引起人类非细菌急性胃肠炎的首要病原体[1]。全世界范围内约1/5的腹泻病例与诺如病毒感染相关,每年约导致20万人死亡[2]。诺如病毒进化变异速度快,每隔2~3年会出现新的流行株引发大流行,其机制[3]主要是:①诺如病毒衣壳蛋白区氨基酸发生突变,其中P区处于高度变异,被认为是免疫识别和与人体组织血型抗原(HBGA)结合的关键部位[4];②基因重组。在过去二十年间诺如病毒GII.4型一直为优势流行株[5],直至2014-2015冬季在多个亚洲地区国家新型GII.P17-GII.17型基因型取代GII.4_Sydney_2012型成为优势流行株[6];但中国CDC报道自2016下半年以来新重组基因型GII.P16-GII.2在诺如病毒暴发病例中检出率超过GII.P17-GII.17及GII.4型,成为我国引起急性胃肠炎暴发的优势基因型[7]。本文收集舟山海岛地区2017年5-11月4起学校发生的急性胃肠炎暴发疫情病例样本,通过扩增其聚合酶-衣壳区(B-C区),近乎完整的聚合酶区(RdRp)及衣壳蛋白区(VP1)基因序列,鉴定引起本轮诺如病毒暴发疫情的基因型别及分子特征,为诺如病毒的防控提供科学的依据。
1 材料与方法
1.1样本来源及处理 2017年5-11月,舟山市定海区两所幼儿园、一所小学及普陀区一所小学共发生4起急性胃肠炎暴发事件,疫情所在的县区疾控共送检40例样本,其中肛拭子21份、呕吐物13份、粪便6份;对固体样本取豌豆大小 (液体取100 μL) 加到1.5 mL EP管中,加入900 μL的pH 7.2的PBS (10 mmol/L) 缓冲液,振荡1 min。然后静置5 min,再以10 000 r / min离心3 min,然后吸取200 μL上清液,使用QIAGEN Reansy Mini Kit试剂盒按照说明书提取病毒核酸。
1.2诺如病毒筛查 使用TAKARA公司One Step PrimeScriptTMRT-PCR Kit试剂盒用于GI型和GII型诺如病毒筛查。其中GI型反应体系25 μL,包括20 μmol/L的JJV1F/JJV1R/JJV1P各0.5 μL,2×RT-PCR buffer 12.5 μL,5 U/μL EX Taq HS 0.5 μL,RT Ezyme Mix II 0.5 μL,双蒸馏水5 μL,RNA 5 μL;GII型反应体系25 μL,包括20 μmol/L的JJV2F/JJV2R/JJV2P各0.5 μL,2×RT-PCR buffer 12.5 μL,5 U/μL EX Taq HS 0.5 μL,RT Ezyme Mix II 0.5 μL,双蒸馏水5 μL,RNA 5 μL,引物探针序列见表1。反应在ABI公司ViiA-7上进行,实验条件设置如下:42 ℃ 10 min反转录,95 ℃ 15 min预变性,45个循环95 ℃ 15 s,60 ℃ 1 min,Ct值≤40判定为阳性。
1.3聚合酶区-衣壳区扩增及测序 将诺如病毒阳性的样本,使用QIAGEN OneStep RT-PCR Kit试剂盒扩增GII型聚合酶区-衣壳区(557 bp)进行分型,反应体系25 μL,包括5xRT-PCR Buffer 5 μL,10 μmol/L dNTP mix 1 μL,RT-PCR Enzyme mix 1 μL,20 U/μL Rnase Inhibitor 1 μL,50 μmol/L MON431/G2SKR各0.5 μL,双蒸馏水11 μL,RNA 5 μL;实验条件设置如下:42 ℃ 30 min反转录;95 ℃ 15 min预变性;40个循环95 ℃ 1 min,50 ℃ 1 min,72 ℃ 1 min;72 ℃ 10 min延伸。取10 μL扩增产物进行1.5 %琼脂糖凝胶电泳,紫外灯下观察结果。阳性扩增产物送至上海生工生物工程有限公司测序。
1.4聚合酶区和衣壳蛋白区扩增及测序 经分型后证实本轮暴发疫情均由诺如病毒GII.P16-GII.2所引起,随机选择其中1份阳性样本使用QIAGEN OneStep RT-PCR Kit 试剂盒扩增近乎完整的RdRp区(1 153 bp)及VP1(1 580 bp),其中扩增RdRp区反应体系25 μL,包括5xRT-PCR Buffer 5 μL,10 μmol/L dNTP mix 1 μL,RT-PCR Enzyme mix 1 μL,20 U/μL Rnase Inhibitor 1 μL,50 μmol/L RdRp-3875F/RdRp-5259R各0.5 μL,双蒸馏水11 μL,RNA 5 μL;扩增VP1区反应体系25 μL,包括5xRT-PCR Buffer 5 μL,10 μmol/L dNTP mix 1 μL,RT-PCR Enzyme mix 1μL,20 U/μL Rnase Inhibitor 1 μL,50 μmol/L VP1-5025F/VP1-6731R各0.5 μL,双蒸馏水11 μL,RNA 5 μL。实验条件设置如下:42 ℃ 30 min反转录;95 ℃ 15 min预变性;35个循环95 ℃ 1 min,58 ℃ 1 min,72 ℃ 2 min;72 ℃ 7 min延伸。取10 μL扩增产物进行1.2 %琼脂糖凝胶电泳,紫外灯下观察结果。阳性扩增产物送至上海生工生物工程有限公司测序。
1.5基因型别鉴定及序列分析 使用MEGA 6.06 Clustal W将扩增的序列进行比对,并采用Neighbor Joining构建系统进化树。使用Simplot软件分析毒株的重组位点,采用200 bp的滑动窗口,20 bp的滑动步伐。使用BioEdit v.7.0.1软件对序列编辑,同源性分析,并将RdRp区及VP1碱基序列翻译氨基酸序列与代表株进行序列比对;同时通过诺如病毒在线分型工具(http://www.rivm.nl/mpf/norovirus/typingtool)鉴定其型别,并与由系进化分析判定的型别进行比较,本文使用的参考序列均来源于NCBI。

表1 诺如病毒分子检测所用引物与探针序列Tab.1 Primer and probe sequence used in molecular detection for norovirus
注:*1-6引物/探针用于 real-time PCR,7-12用于常规RT-PCR
2 结 果
2.1流行概况 2017年5-11月舟山市海岛地区定海区两所幼儿园与一所小学、普陀区一所小学分别发生急性胃肠炎暴发疫情,总病例数69例,暴发流行时间最短4 d,最长10 d;疫情呈明显聚集性多发生于同一宿舍、班级及相邻班级。病例年龄组在3~10岁,临床表现以恶心、呕吐为主伴随不同程度发热、腹泻、腹痛,无重症病例及死亡病例。结合流行病学调查及实验室检测结果判定该起疫情排除水源性和食源性传播的可能性,认为是由诺如病毒感染引起的人传人暴发事件。
2.2诺如病毒筛查结果 对送检的40份样本进行诺如病毒GI型和GII型荧光定量检测,其中阳性样本27例,均为诺如病毒GII型,阳性检出率67.50%,见表2。
2.3聚合酶区-衣壳区扩增结果及序列分析 聚合酶区-衣壳区扩增产物经琼脂糖凝胶电泳分析,27份样本均在557 bp出现特异性条带。送至上海生工生物工程公司进行序列测定,均测序成功,经诺如病毒在线分型工具判定均为GII.P16-GII.2型,见表2。系统进化分析结果显示GII.P16-GII.2型27份,与在线分型工具所得结果一致,各个毒株间序列高度同源,相似性为98.8%~100%,见图1。

表2 4起暴发疫情诺如病毒分子检测结果Tab.2 The molecular detection results of norovirus in four acute gastroenteritis outbreaks
2.4聚合酶及衣壳蛋白区扩增结果及序列分析 聚合酶区和衣壳蛋白区扩增产物经琼脂糖凝胶电泳分析,均出现特异性条带。送至上海生工生物工程公司进行序列测定,测序成功后经诺如病毒在线分型工具判定为GII.P16与GII.2型。将所得序列提交至NCBI,获得序列号:MH057762。Simplot软件重组分析显示,重组位点位于ORF1-ORF2连接区处1 120-1 122 bp(AY134748,5 052-5 053 bp),见图2。 经系统进化结果可见,2016-2017年新重组型GII.P16型及GII.2型均来源于2010-2012年日本的GII.P16-GII.2毒株,本次舟山海岛地区幼儿园中流行暴发的GII.P16型隶属于GII.P16c簇与2016江苏的毒株同源性最高,为99.7%,GII.2型隶属于GII.2c簇与2016年北京的毒株同源性最高,为99.9%,见图3,4。将本次分离出来的新重组毒株GIIP.16-GII.2与代表株进行RdRp区及VP1区氨基酸序列比对,均发现多个位点发生突变,在RdRp区D173E、S293T、V332I、K357Q及T360A位点的突变仅发生在2016-2017年重组株的GII.P16c簇中;与GII.2代表株VP1区氨基酸序列相比,GII.2 a,b,c 三簇在P区均有多个位点突变,在潜在抗原表位及与人体HBGA结合的位点周围共12个氨基酸位点发生突变,2016-2017年的GII.2c簇中并不存在特异性突变。见表3,4。

注:诺如病毒命名规则按诺如病毒/基因型/宿主/国家/年份/基因亚型/分离地点/序号[8]图1 27株诺如病毒聚合酶区-衣壳区序列系统进化分析结果Fig.1 Phylogenetic analysis on partial RdRp and capsid protein sequences of 27 norovirus strains in Zhoushan Islands,2017

图2 Simplot分析2017年舟山海岛GII.P16-GII.2重组位点结果Fig.2 The recombination breakpoint result of GII.P16-GII.2,Zhoushan Islands,2017

图3 GII.P16型RNA聚合酶区序列系统进化分析结果Fig.3 Phylogenetic analysis results of GII.P16 RdRp sequences

图4 GII.2型VP1区序列系统进化分析结果Fig.4 Phylogenetic analysis results of GII.2 VP1 sequence

表3 舟山市海岛地区GII.P16型诺如病毒RdRp区氨基酸序列变异结果Tab.3 The amino acid sequence mutation of GII.P16 RdRp region in Zhoushan Islands
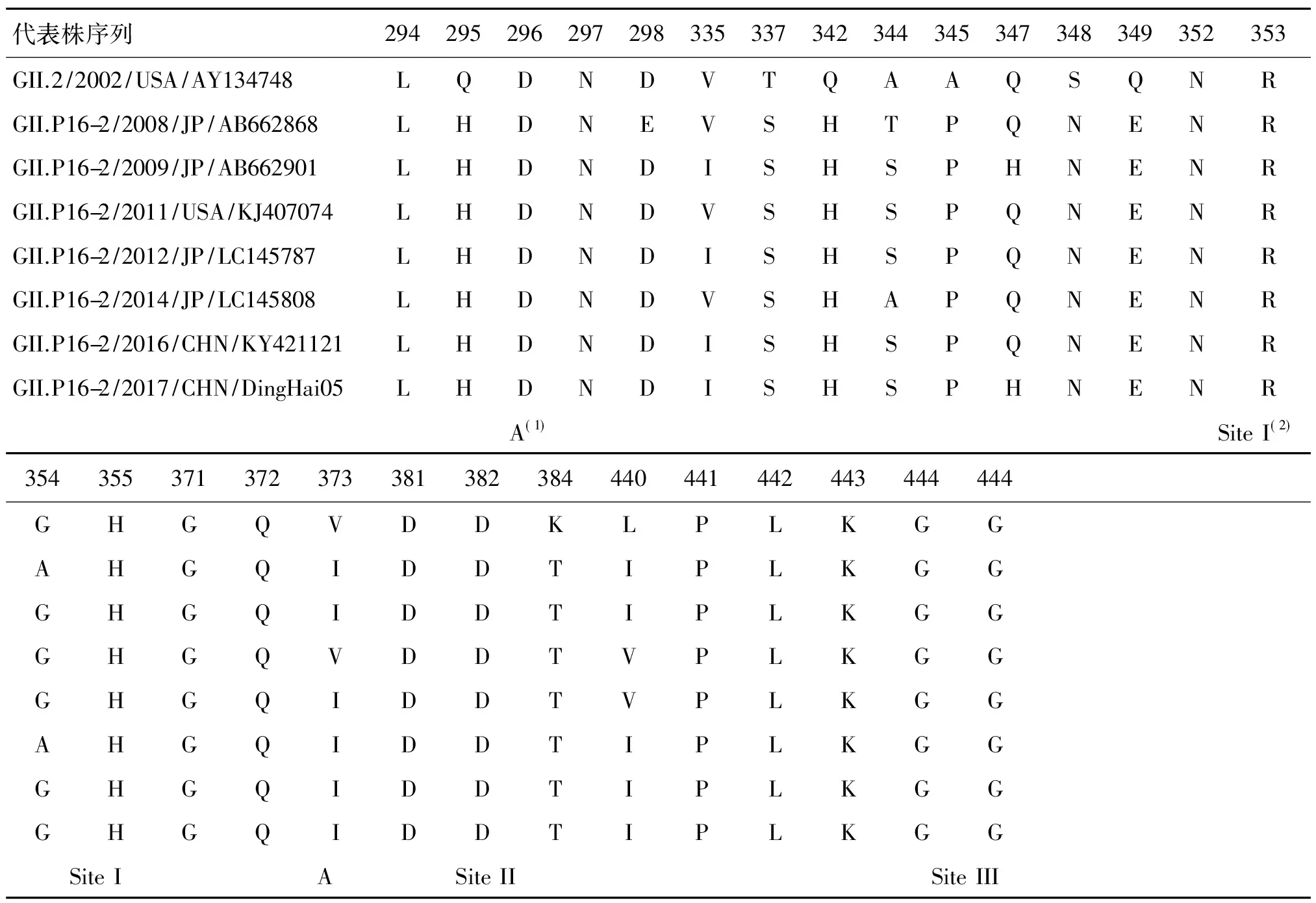
表4 舟山市海岛地区GII.2型诺如病毒衣壳蛋白区氨基酸序列变异结果Tab.4 The amino acid sequence mutation of GII.2 VP1 region in Zhoushan Islands
注:(1)由GII.4型提示可能潜在的诺如病毒A抗原表位位点(294,296-298,368,372)[9]
(2)GII.2与人体HBGA受体结合的3个位点[10]
3 讨 论
诺如病毒为杯状病毒科诺如病毒属,为单链正股RNA病毒,全长约7.5-7.7 kb,整个基因组可分为ORF1、ORF2、ORF3三个开放读码框(ORFs),分别编码包含RNA聚合酶(RdRp)的非结构蛋白,主要衣壳蛋白(VP1)及次要衣壳蛋白(VP2)[11]。基于RdRp区和VP1基因序列的差异可将诺如病毒分为GI-GVII 7个基因群,GI,GII,GIV可感染人类[12]。2017年5-11月舟山市海岛地区发生4起由诺如病毒GII.P16-GII.2型引起的急性胃肠炎暴发疫情,且都发生在低年龄人群学校场所。中国CDC一篇文献报道,2016年我国大陆由诺如病毒引起的急性胃肠炎暴发中新重组的GII.P16-GII.2型检出率为79%[7];同期我国香港及台湾、德国、日本、法国等报道自2016年以来GII.P16-GII.2型诺如病毒引起的暴发疫情显著增加,且与本区域类似,大多数暴发疫情是发生在幼托,幼儿园,小学等场所[13-17];吴冰珊等认为GII.P16-GII.2型类似于GII.4型,具有在局部人群的传播能力[18];但GII.4型多暴发于养老院等长期的健康护理机构[19];传播方式倾向于人与人接触传播[20],GII.P16-GII.2传播模式是否类似于GII.4型还需进一步验证;本次舟山海岛地区分离出来的毒株GII.P16,GII.2分别与同处沿海地区江苏省,北京市2016年报道的GII.P16及GII.2型毒株同源性最高,此前有文献报道,对1999-2011年我国的诺如病毒重组株分析,多数型别均分布在沿海地区,且推断受诺如病毒污染海产品的广泛销售有助于诺如病毒基因型的传播与分布[21]。
舟山海岛地区分离出来的GII.P16-GII.2型毒株,扩增近乎完整的RdRp区及VP1区,通过与代表株序列比对,在RdRp区D173E、S293T、V332I、K357Q及T360A位点的突变仅发生在2016-2017年重组株GII.P16c簇中;与GII.2代表株VP1区氨基酸序列相比,GII.2 a,b,c 三簇在P区均有多个位点突变,但在2016-2017年的GII.2c簇中并不存在特异性位点突变。在GII.4型及GII.P17型的进化过程中,P区氨基酸的变异往往对新流行株的出现,及其抗原性的转变与易感人群范围的扩大有关键性的作用,使得新流行株能够逃避人群免疫力而引起广泛的流行[22-23],而本次新重组GII.P16-GII.2型VP1区序列分析结果提示P区位点的变异并不是促使本轮新重组株GII.P16-GII.2暴发流行的关键因素;采用Simplot软件对本次分离的毒株的重组位点进行分析,结果显示重组位点位于ORF1/ORF2连接处1 120-1 122 bp(AY134748,5 052-5 053 bp),该区域为诺如病毒发生基因重组的主要区域[24],基因重组是诺如病毒的进化重要机制之一,往往发生于不同亚型的混合感染病例中,衣壳蛋白区结合不同的聚合酶区,能够改变病毒的复制与传播能力,或者产生新的诺如病毒亚型,造成抗原性漂变,从而能够逃避人群免疫力,引发大范围的暴发流行[25]。值得注意的是近年GII.P16的重组株不断被发现,表明GII.P16型RdRp区已进化具备结合多种型别衣壳蛋白区的能力,对最近出现的重组株GII.P16-GII.2及GII.P16-GII.4_Sydney2012型RdRp区分析发现有5个氨基酸位点特异性突变,并且此5个位点靠近RdRp功能区[26],有学者推测GII.P16型RdRp区这些位点的突变改变了新重组型病毒的传播能力,从而使其能够广泛的流行[27];由于目前对于GII.P16-GII.2认识尚还不全面,这些氨基酸位点突变对于GII.P16-GII.2型的流行的贡献还待进一步确认。
舟山市海岛地区在本轮暴发疫情中首次检测到了诺如病毒新重组型GII.P16-GII.2,提示在今后的监测工作中开展诺如病毒的RdRp及VP1区序列检测、监控本区域内诺如病毒重组情况及特殊氨基酸位点的改变,以期及时识别新出现的变异株,同时加强监测力度,明确该重组型别流行特征、趋势及是否存在特异性临床特征及易感人群的改变,为本区域诺如病毒的防控提供科学的依据。
