西番莲解剖结构特征及低温胁迫下叶片结构与抗寒性的关系
2018-12-04李安定郝自远张洪亮张丽敏蔡国俊
滕 尧,李安定,* ,郝自远,张洪亮,张丽敏,蔡国俊
(1.贵州科学院 贵州省山地资源研究所,贵州 贵阳550001;2.南京林业大学 林学院,江苏 南京210037;3.贵州科学院,贵州 贵阳550001)
西番莲(Passiflora caerulea L.)是原产于热带地区的草本或常绿攀缘木质藤本多年生植物,属被子植物门双子叶植物纲西番莲科(Passifloraceae)西番莲属(Passiflora L.)[1]。目前,用于食用或加工的西番莲品种果实的果汁含量为30% ~40%,含130种以上的芳香物质,故又名“百香果”。我国于1913年由菲律宾引进种植西番莲,主要引种到温度较高的沿海地区[2],主要栽培地区包括台湾、海南、福建、广东、广西和云南。目前,各地种植的西番莲品种主要是紫果西番莲(Passiflora edulis Sims)和黄果西番莲(Passiflora edulis var flovioarpa Degener)以及二者的杂交品种[3],代表品种有台农1号、华阳511、紫香1号等。
贵州省属于喀斯特山地,石漠化问题由来已久,而西番莲常绿、攀缘藤本、生长迅速且能产生经济效益等特点,使其成为喀斯特地区石漠化防治的理想栽培作物[4]。贵州省年均气温16℃左右,除黔南州的少数地区外,其他地区冬季极端低温都在0℃左右,甚至更低,并持续数周,这会导致西番莲死亡[5]。近10年来,贵州科学院山地资源研究所喀斯特山地高效特色藤本植物研究团队,在喀斯特山地进行了西番莲引种试验及高效栽培研究,培育出了1个具有较强抗寒性的品种(平塘1号)。另外,贵州不同气候地点经多年驯化选育出4个优良基因型(黔乡香1、2、3、4号),其果实外型大小、颜色和风味及抗寒性与其他品种均存在一定差异。植物叶片对寒冷较为敏感,在低温胁迫下其形态结构和生理生化适应性调整较为活跃[6-8]。本文研究了平塘1号、黔乡香4个基因型与紫香1号一年生扦插苗的生理结构差异,以及2个品种叶片结构在抗寒性上的差异,旨在为西番莲抗寒新品种的选育及推广应用提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于贵州省黔南州平塘县克度镇光明村(106°48'19″E,25°43'37″N),海拔 853 m,年平均气温16.3℃,年降雨量1 400 mm左右。雨季为5—10月,占全年降雨量的80%。属亚热带季风气候,土壤为山地黄壤。
1.2 材料
供试材料为平塘1号、黔乡香1号、黔乡香2号、黔乡香3号、黔乡香4号和紫香1号的一年生扦插苗标准株。常规解剖材料包括2级侧根的根尖成熟区、茎上进行花芽分化的2级侧枝、2级侧枝顶端当年生叶片、正常开放的成熟花朵、成熟的果实及其种子。低温胁迫处理的解剖材料为温室大棚中培育的长势正常、无病虫害的一年生平塘1号和紫香1号整株扦插苗。
1.3 方法
解剖观察采用石蜡切片法进行制片,具体制片过程参照《植物显微制片技术实验指导书》,在显微镜下观察并拍照。
低温胁迫处理选择本地适生、抗寒性强的平塘1号和果实品质好但抗寒能力较弱的紫香1号为材料。低温胁迫温度梯度设定参考试验地克度镇历年气象统计数据,尽可能还原试验地的温度、光照、湿度等气候条件。试验地历年观测结果表明,紫香1号的极限耐寒温度为4~8℃(个体存在差异),长期处于该温度范围内亦会发生冷害。根据文献[5]和[9]的结果,将本次研究的低温胁迫梯度设置为5.0、2.5、0、-2.5℃,低温胁迫时间设置6、12、15和24 h,并在前述温度设置下进行预处理。本研究中,紫香1号5.0℃处理24 h未出现明显的冷害症状,2.5℃处理24 h后开始出现轻微冷害影响;0℃处理时,其叶片在13~15 h开始出现冻害;-2.5℃处理,其叶片在10~12 h出现明显冻害。-2.5℃处理24 h后平塘1号未发现明显冻害症状。因此,本研究将低温胁迫处理时间设置为12 h。
按上述温度梯度及时间处理完成后,在显微镜下观察和拍照,测量并记录叶片的各个结构,所得数据为12个视野内的平均值。所测叶片解剖结构包括:叶片长度、叶片宽度、叶片厚度、叶脉厚度、蜡质层厚度、叶脉维管束宽度、叶脉维管束长度、海绵组织和栅栏组织,共9项,并计算叶片组织结构紧密度、叶片组织疏松度、叶脉突起度。
叶片组织结构紧密度(cell tense ratio,CTR,%)=栅栏组织厚度/叶片厚度×100;
叶片组织疏松度(spongy ratio,SR,%)=海绵组织厚度/叶片厚度×100;
叶脉突起度(vein protuberantdegree,VPD,%)=叶脉厚度/叶片厚度×100。
云天化示范田与其它品牌对照田相比,示范田的肥料成本略高,但示范田的产量高于对照田,品质更好收购价高,示范田纯收入要高于对照田,实现亩增效益912.4元。这表明,使用云天化系列化肥可以显著提高该地区樱桃种植户的种植效益。
1.4 数据分析
使用Excel进行原始数据的记录及整理,图片整合及分辨率调整使用Photoshop CC(14.0)完成,数据运算分析使用SPSS 19.0。
2 结果与分析
2.1 西番莲不同器官解剖结构
2.1.1 根
对西番莲根的横切、纵切片进行观察和对比,未发现某个品种或基因型的西番莲根在某一个组织结构上存在明显的差异。从横切图(图1)可以看出,西番莲根的组织结构包括次生韧皮部、形成层、次生木质部和侧根。次生木质部占据根横切面积的2/3左右,其内具有各种直径的导管,排列分布不规则;形成层介于次生木质部和次生韧皮部之间,结构明显;次生韧皮部中筛管、伴胞和薄壁组织均较发达。纵切片(图2)中也未发现明显的组织结构差异。
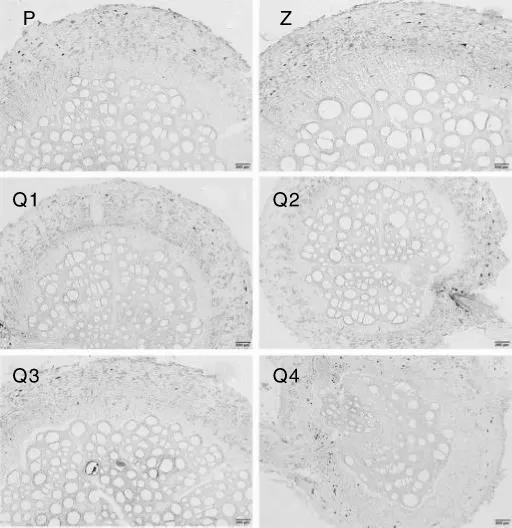
图1 西番莲根结构解剖横切图Fig.1 Cross cutting anatomic structure pictures of Passiflora edulis roots
2.1.2 茎
在外观形态上,不同品种和基因型西番莲茎的主干上都有较明显的节间,进行花芽分化的分枝上则没有节间。茎的解剖横切图(图3)中未发现不同品种和基因型西番莲有明显的组织结构差异,茎的形态结构清晰,主要结构包括表皮、皮层、维管束(初生韧皮部、束中形成层和初生木质部)、髓射线和气腔;表皮细胞呈砖形或圆形,没有细胞间隙,排列紧密;皮层中细胞排列疏松,有间隙;维管束各层结构明显,细胞排列紧密,形状不规则;维管束之间具有比较明显的束间薄壁组织,即髓射线;气腔呈椭圆形,位于结构中央,占整个茎横截面积的1/2左右。茎的纵切图(图4)中也未发现明显的组织结构差异。
2.1.3 叶
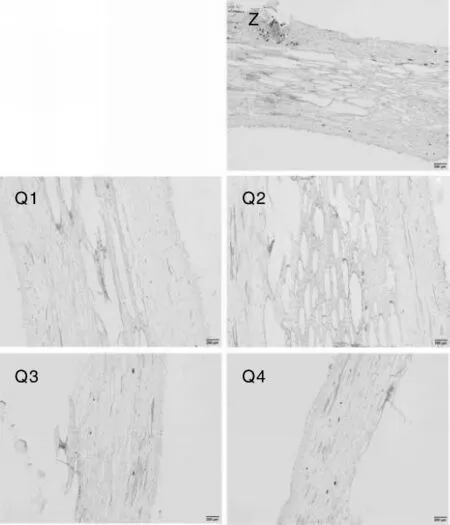
图2 西番莲根结构解剖纵切图Fig.2 Rip cutting anatomic structure pictures of Passiflora edulis roots
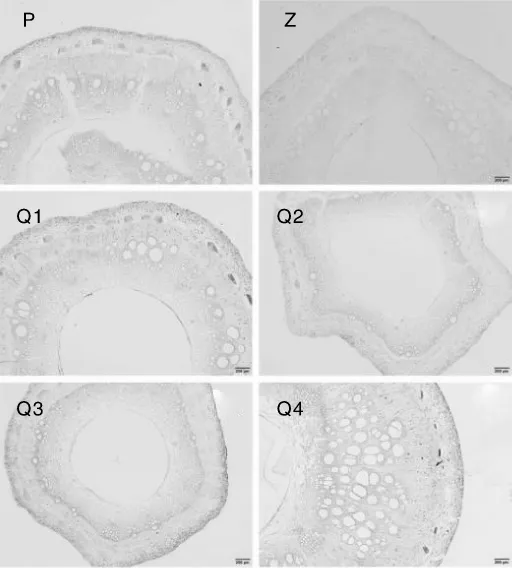
图3 西番莲茎的结构解剖横切图Fig.3 Cross cutting anatomic structure pictures of Passiflora edulis stems
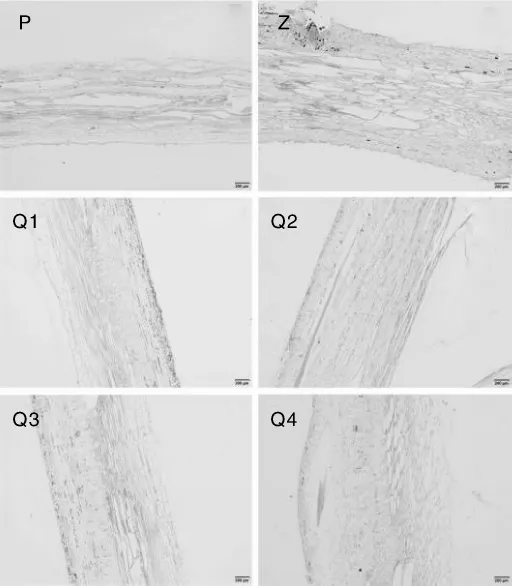
图4 西番莲茎的结构解剖纵切图Fig.4 Rip cutting anatomic structure pictures of Passiflora edulis stems
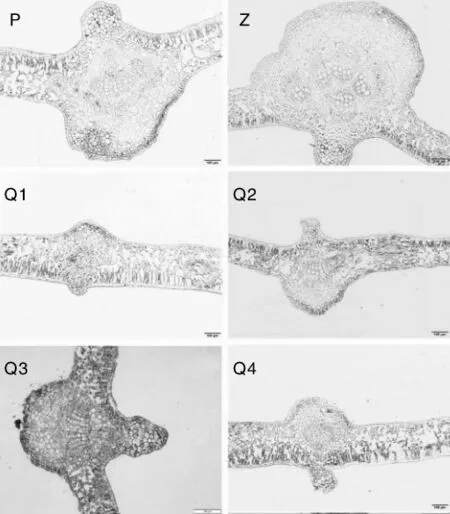
图5 西番莲叶的主脉结构解剖图Fig.5 Anatomic structure of leaf veins of Passiflora edulis leaves
不同品种和基因型西番莲的叶片外观形态结构基本一致,仅大小不同。在叶主脉的解剖结构切片图中,可观察到叶片由3部分组成:叶脉、表皮和叶肉。叶脉包括维管束、薄壁组织和厚角组织,维管束木质部导管发达。叶主脉解剖结果(图5)显示,与紫香1号相比,平塘1号和黔乡香4个基因型木质部导管在结构上有明显的区别,它们的上表皮厚角组织下方均有一簇较小的木质部,在小木质部下方有一簇较大的木质部。平塘1号和黔乡香4个基因型的大木质部导管呈整齐排列的束状,向下以辐射状展开,紫香1号的大木质部导管呈聚合式的不规则团状围绕在小木质部周围。表皮由一层扁平状细胞组成,排列紧密,无胞间隙,上表皮细胞外切向壁覆有较厚的蜡质层,下表皮有气孔。叶肉包括位于上、下表皮之间的栅栏组织和海绵组织,靠近叶片上表皮细胞处为栅栏组织,细胞长柱形,排列较紧密,其长轴垂直于上表皮,成栅栏状,细胞内清晰可见叶绿体;栅栏组织下方为海绵组织,细胞形状不规则,排列较疏松,有发达的胞间隙,细胞内有较多叶绿体(图6)。
2个品种和黔乡香4个基因型西番莲花的外观形态仅在花柱的紫色斑点上存在差别。紫香1号花柱的紫色斑点数量较多且分布密集,花柱大部分被紫色斑点覆盖;黔乡香4个基因型和平塘1号的花柱上的紫色斑点数量较少,零星分布于柱头附近。西番莲花包含花丝、雌蕊和雄蕊,未发现不同品种或基因型西番莲花在组织结构上有明显的差异,为展示方便,拍照时选取整体和不同局部特征结构进行拍照(图7)。花丝呈圆形、长椭圆形,以游离状分布于雌雄蕊周围;雄蕊的花丝短于雌蕊的花柱,花药的花粉囊呈圆形,其中含有许多花粉母细胞;雌蕊由外向内依次可观察到子房壁、胎座、胚珠和子房室,胚珠为不规则椭圆形,包含珠柄、珠被、胚囊和珠心。

图6 西番莲叶肉结构解剖图Fig.6 Anatomic structure of mesophyll of Passiflora edulis leaves
2.1.5 果实
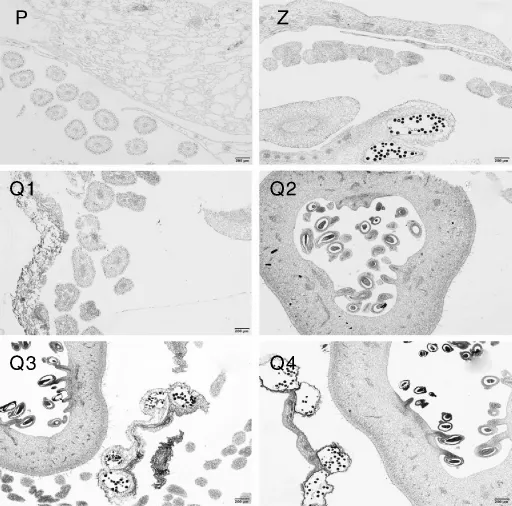
图7 西番莲花的结构解剖图Fig.7 Anatomic structure pictures of Passiflora edulis flowers
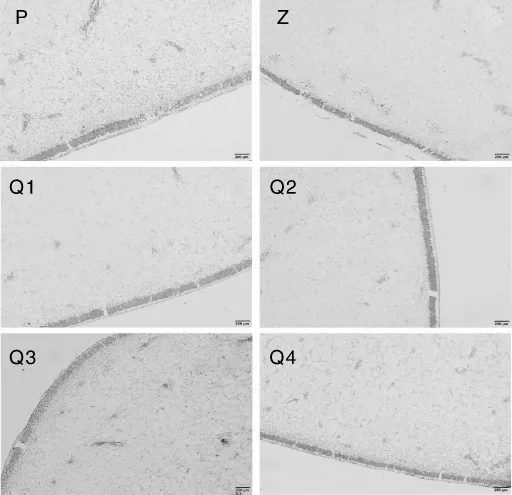
图8 西番莲果实的结构解剖图Fig.8 Anatomic structure pictures of Passiflora edulis fruits
不同品种和基因型西番莲果实在形状、大小、果皮颜色、果皮表面斑点、海绵层厚度、胎座颜色、果汁成色及风味上均有较明显的差异,但果实的形态解剖结构未发现明显差异(图8)。果实的解剖结构包括果皮、海绵层、胎座、内果皮和种子。最外层为表皮细胞,表皮细胞下有一层排列紧密的石细胞,石细胞内为基本组织,基本组织的外层是海绵层。不同品种和基因型西番莲的海绵层均为白色,但海绵层的厚度不同。海绵层上有许多连接种子的脊,即胎座,平塘1号和黔乡香4个基因型的胎座呈粉红色,紫香1号的胎座呈白色。胎座连接着内果皮,内果皮内部包裹着种子,成熟果实的内果皮中有大量果汁,即西番莲的主要食用部分。
2.1.6 种子
不同品种和基因型西番莲的种子在成熟时均为阔卵形、黑色,种子大小和千粒重也基本一致。根据植物种子的结构特征,完整的种子结构应包括胚(胚根、胚芽、胚轴及子叶)、胚乳和种皮3部分,双子叶植物种子的胚乳在种子休眠前基本完全被胚胎吸收[10]。由于样本采集时间为11月,受低温影响果实并未完全发育成熟,导致未成熟的种子在制片过程中达不到理想效果。在可进行解剖观察的西番莲种子(图9-Q1)中可看到,种子处于生长发育阶段,可观察到种皮经染色后呈波浪形包裹在外围,种皮内有大量胚乳,子叶处于生长发育阶段,还未形成明显的双子叶形态,胚芽还未成形,胚轴和胚根已具有一定形态,但均未发育成熟(图9)。
2.2 低温胁迫下西番莲叶片的解剖结构特征及其与抗寒性的关系
2.2.1 叶片的解剖结构特征
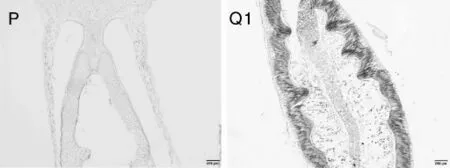
图9 平塘1号及黔乡香1号西番莲种子的结构解剖图Fig.9 Anatomic structure pictures of Ping Tang 1 and Qian Xiang Xiang 1 seeds
植物生理结构上起主要抗寒作用的器官是叶片[8,11-12]。在平塘 1 号和紫香 1 号的叶主脉解剖结构中发现,随着温度的不断降低,木质部维管束的形态并未发生明显的适应性改变。二者的栅栏组织在不同低温下均呈排列有序的状态,但在排列致密程度上平塘1号大于紫香1号,在温度下降时这一特点更加明显,平塘1号的栅栏组织明显长于紫香1号(图10)。二者的海绵组织均呈疏松和零散分布,随着胁迫温度的不断降低,它们的海绵组织细胞逐渐变得致密,厚度也不断增加。温度在0℃及以下时,海绵组织和栅栏组织的结构分界不清晰。
2.2.2 叶片结构与抗寒性的关系
如表1所示,5.0℃处理12 h,2个品种的一年生扦插苗叶片组织结构表现出了不同的适应性变化,二者在叶片长度、宽度及厚度上均表现出了显著差异。受叶片大小的影响,平塘1号的叶脉维管束长度极显著小于紫香1号,宽度显著小于紫香1号。二者蜡质层厚度、叶脉厚度和栅栏组织差异不显著,海绵组织、组织结构紧密度、疏松度和叶脉突起度差异均显著。平塘1号叶片较小而叶脉更厚的结构特征,可能是其在面对低温度胁迫时组织结构发生适应性调整的前提条件。
2.5℃处理12 h后,平塘1号和紫香1号的叶片长度、宽度及厚度,以及叶脉厚度和维管束长度均呈现显著性差异,蜡质层厚度差异不显著。与5.0℃处理相比,2.5℃处理下2个品种海绵组织的厚度均有一定程度的增加,增加后二者差异不显著。平塘1号在2.5℃胁迫下适应性更强,其通过调整栅栏组织的长度来提高组织结构紧密度,以达到更好的抗寒效果。平塘1号经驯化后,为了抵抗寒冷而拥有更厚的叶脉,这也使得二者在2.5℃下组织结构紧密度和叶脉突起度呈现极显著差异。
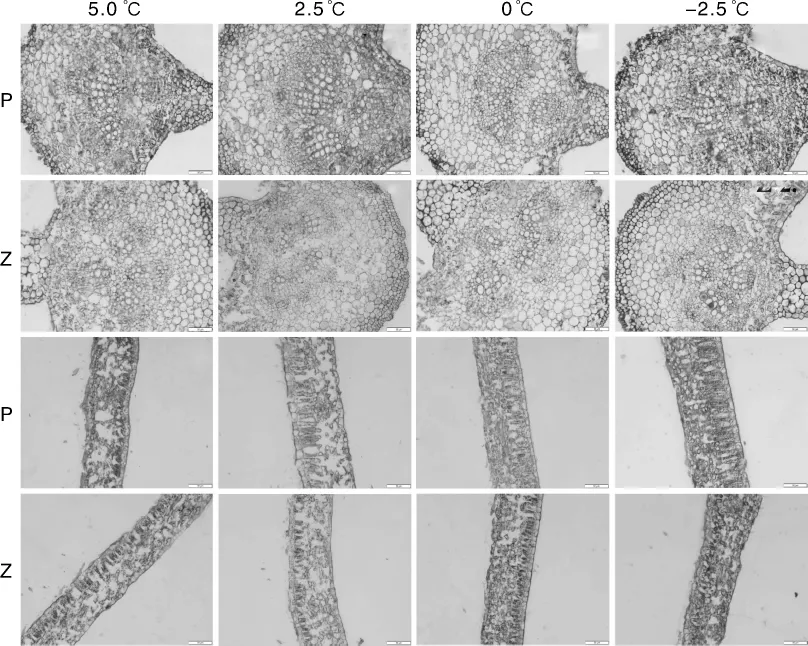
图10 西番莲叶片在不同低温胁迫下的叶脉及叶肉解剖结构图Fig.10 Anatomical structure of Passiflora edulis leaf vein and mesophyll under different low temperature

表1 5.0、2.5、0和-2.5℃处理12 h后2个品种西番莲叶肉组织的结构比较Table 1 Comparison of the mesophyllous structure of 2 Passiflora edulis varieties at 5.0,2.5,0 and -2.5℃ after 12 h treatment
0℃处理12 h后,2个品种叶片长度、宽度及厚度,以及叶脉厚度依旧呈现显著性差异,维管束长度和蜡质层厚度差异不显著,但维管束宽度却呈现出了极显著差异。平塘1号海绵组织厚度极显著小于紫香1号,但栅栏组织长度极显著大于紫香1号,说明抗寒品种对栅栏组织结构的适应性调整是抗寒的关键。平塘1号的叶片厚度显著小于紫香1号,所以其组织结构疏松度反而较大,但这并不是决定平塘1号抗寒性的主要因素。推测由叶脉厚度和栅栏组织长度差异导致的组织紧密度和叶脉突起度差异,是在形态结构上决定品种抗寒性大小的关键因素。
-2.5℃处理12 h后,2个品种的叶片长度和宽度差异不显著,推测是因为-2.5℃已经达到了紫香1号的半致死温度[5],经过12 h处理,紫香1号的叶片已经产生一定程度的萎蔫现象,叶片变色并产生褶皱和卷曲,这也间接导致了叶脉厚度、蜡质层厚度、维管束长度及宽度发生不同程度变化。与平塘1号进行显著性检验时,除叶片厚度外,上述指标与5.0、2.5、0℃处理的差异较大,而平塘1号的叶片外观并未发生改变,组织结构测量数据亦显示平塘1号未受冻害,依然能通过调整自身组织结构来适应寒冷,证明紫香1号的半致死温度对平塘1号未造成实质性损害,平塘1号的抗寒能力大于紫香1号。
综上,西番莲叶片的抗寒性主要取决于叶片长度、叶片宽度、叶片厚度、叶脉厚度与栅栏组织长度,海绵组织厚度对抗寒性也有一定影响,蜡质层厚度对西番莲的抗寒性几乎没有影响。维管束的长度与宽度在不同低温胁迫处理时未表现出一定的规律性,所以,西番莲不同品种维管束结构差异对抗寒性是否有影响还需做进一步研究。
3 讨论
3.1 叶片解剖结构特征与其抗寒性的关系
抗寒性是植物对寒冷冻害长期适应的一种遗传特性,抗寒性主要取决于植物自身的形态结构特征变化和植物内部的生理生化变化,也取决于植物所处的外界环境条件。植物的叶片是最先感受寒冷且对寒冷特别敏感的器官[8,11-12]。研究认为,植物的抗寒性与叶片长短及厚度呈显著性负相关,即叶片长宽越大、叶片越厚的品种,其抗寒能力越弱,反之则强;与组织结构紧密度和叶脉突起度呈显著性正相关,即组织结构紧密度和叶脉突起度越大的品种,其抗寒性也越好[13]。受到低温胁迫时,植物组织会调整自身的形态结构以适应低温带来的影响。如本研究中平塘1号的栅栏组织平均长度及平均叶脉厚度随着温度的降低逐渐增大,以提高组织结构紧密度和叶脉突起度,从而提高抗寒性;而紫香1号的组织适应调整不如平塘1号明显,所以其抗寒性相对较弱,但这并不能在本质上决定该品种的抗寒能力,还需参考其低温胁迫下的生理生化指标变化、田间种植情况等[12-14]。不同温度处理后,平塘1号的叶片长宽更小、叶片厚度更薄,外在形态上符合抗寒条件的特征,其叶脉厚度更厚、栅栏组织长度更长,表现出了更好的组织结构紧密度和叶脉突起度,也满足抗寒性的结构特征。叶脉维管束形态、长度与宽度的差异可能会导致植物在水分及养料供应上存在区别[15-16],这可能是两个品种在抗寒性上存在差别的原因之一。但也有研究表明,叶片的叶脉维管束面积比率对植株的抗寒性基本没有影响[7],所以对不同品种或基因型的西番莲而言,叶脉维管束形态和大小的差异是否会对植株的抗寒性产生影响有待进一步研究。二者的组织结构疏松度虽然有显著性差异,但并不是影响品种间抗寒性差异的主要因素,其原因是平塘1号的海绵组织厚度和叶片厚度均小于紫香1号,比例换算时平塘1号的组织结构疏松度反而大于紫香1号,但就植株的叶片而言,更大更厚的叶片表示其叶肉细胞就有更大的表面积进行光合作用,这很可能是紫香1号在产量和果实品质上均优于平塘1号的主要原因。
3.2 西番莲抗寒性的综合指标
抗寒性是从植物的形态结构、水分生理生态特征及生理生化反应到组织细胞、光合器官乃至原生质结构特点的综合反映[13]。植物的抗寒性是由不同层面的综合因素影响决定的,不能简单地从一个或几个指标来反映其抗寒性。许多研究已证明[7,17-18],多个指标的综合评价运用,才能较客观地反映植物的抗寒能力强弱。本研究中2个不同品种西番莲在低温胁迫下的叶片形态结构说明,叶片在低温环境下组织结构上的适应性调整是该品种抗寒能力的主要反映指标之一,也能在一定程度上代表西番莲抗寒能力的强弱。但不能因此就对不同品种西番莲的抗寒能力强弱下定论,因为抗寒性的强弱与许多因素有关,如植物生理生化的应激变化、植物根系是否发达以及田间病虫害等。本研究只对西番莲叶片形态结构与其抗寒性的关系进行了分析,如果要对西番莲的抗寒性进行综合评价,需对其在低温胁迫下的生理生化特征变化、抗寒基因的表达等影响因素开展进一步研究。
4 结论
在外形上,平塘1号、4个黔乡香系列优良基因型与紫香1号在各自的果实形态与风味口感上存在差异,雌蕊柱头上的紫色斑点数量与密度也存在差异;在形态学解剖结构上,平塘1号、黔乡香4个基因型与紫香1号的叶片木质部导管有明显差异,其他组织结构在本次制片观察中未发现明显差异。
叶片的大小、厚度及栅/海比是影响西番莲植株抗寒性的主要形态结构指标,叶片较小、较薄的品种,其抗寒性较强,反之则较弱;植物的抗寒性与叶片细胞的结构紧密度和叶脉突起度呈正相关,栅栏组织所占比例越大、叶脉厚度越厚,叶片的组织结构紧密度及叶脉突起度就越大,品种的抗寒性越强,反之则弱。低温胁迫下平塘1号和紫香1号的转录组测序结果表明,多个关键抗寒基因在平塘1号中的表达水平均高于紫香1号,这表明平塘1号的耐寒能力可能高于紫香1号[19]。综合本次试验结果、田间观测结果及之前的研究结果来看[5,9,19],2 个品种及黔乡香 4 个优良基因型的西番莲抗寒性强弱应为:平塘1号>黔乡香系列>紫香1号。
