不同砧木对马瑟兰葡萄抗旱性的影响
2018-11-28任天赐徐美隆
施 明,刘 静,仝 倩,任天赐,徐美隆
(1.种苗生物工程国家重点实验室,银川 750004;2.宁夏大学生命科学学院,银川 750021;3.宁夏大学葡萄酒学院,银川 750021)
世界上葡萄栽培的国家在100多年前由于受到根瘤蚜的危害几乎都采用了抗根瘤蚜的砧木进行嫁接栽培,又存在葡萄生长的不利生态因素如高寒、干旱、盐碱、高湿等,葡萄砧木的研究和嫁接技术迅速发展[1-4]。据调查统计,欧洲国家如法国、德国等葡萄嫁接栽培已超过了葡萄总面积的95%[5]。
查阅资料发现我国有关葡萄砧穗互作机理的研究起步较晚,有关葡萄砧木品种抗旱性的研究报道较多[6-9],但关于葡萄砧木对接穗的生长势、结果状况、产量以及抗逆性等方面还缺乏系统的研究。已有的研究表明:葡萄砧木对接穗的生长和结果特性等都有明显的影响,可有效提高接穗的抗病性和抗逆性,缓解葡萄生产中的病虫害与逆境伤害问题,并且受到品种特性、地域条件、气候环境等多方面因素的影响[10-12],因此,开展砧穗互作机理研究很有针对性。本试验选择3个砧木,嫁接马瑟兰与马瑟兰自嫁接苗形成4个嫁接组合,研究4个嫁接组合在干旱胁迫下的光合及生长情况,为西北干旱半干旱区葡萄嫁接栽培提供理论支撑。
1 材料与方法
1.1 试验材料
试验材料为3个砧木3309A、5BB、1103P,马瑟兰自嫁接苗为对照。3个砧木及马瑟兰的枝条于2月底电热温床催根营养袋扦插,5月底进行了绿枝嫁接,培养成嫁接苗。
1.2 试验方法
本试验采用盆栽法,于6月底将上述营养袋嫁接苗移栽至盆高300 mm,盆口径280 mm,盆底径220 mm的花盆中,盆中装15 kg营养土(黄土∶草炭∶珍珠岩=2∶1∶0.5,再加入总体积8%的有机肥),每盆一株,移栽后浇透水,之后正常管理。经培养60 d后,从8月下旬开始,在抗旱棚内进行干旱胁迫。试验设置处理组5株,对照组5株,重复3次,随机区组排列,采用随时间推移的自然水分胁迫,对照组正常浇水。于第0、7、14、21、28 d测定光合及相应的生长、生理指标用于统计分析。
1.3 测定指标
(1)株高和径粗调查:胁迫前和胁迫后测定各处理株高和径粗,株高用卷尺测量,径粗利用游标卡尺测量嫁接口上下8 cm处主干粗度。
(2)光合作用参数测定:选取各处理健康枝条第6片功能叶,采用GFS-3000光合仪测定,于晴天上午9-10点测定光合参数,包括叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr),每个处理测定9株。
(3)叶绿素和质膜透性测定:叶绿素含量采用乙醇浸提比色法测定;质膜透性采用电导率法测定。
(4)旱害指数的测定:以干旱处理条件下植株叶片形态变化为基础,于干旱胁迫处理后调查植株的旱害症状,确定旱害级别。划分为5级:1级:植株叶片正常;2级:植株1/5以下叶片有轻度的失绿症状;3级:植株1/3以下叶片发黄;4级:植株1/2以下叶片发黄:5级:植株4/5以上叶片发黄。
旱害指数=∑(旱害级数×相应旱害级植株)/
(总株数×旱害最高极值)×100
(1)
1.4 数据分析
采用Excel 2007和SPSS19.0统计软件进行数据处理和方差分析。图表中数据为“平均数±标准差”。
2 结果与分析
2.1 干旱胁迫对不同嫁接组合叶片质膜透性的影响
植物细胞膜的透性在干旱胁迫条件下变化越大,细胞受伤害就越重,对应的葡萄品种的抗旱性就越弱[13]。由图1可知,各嫁接组合处理在干旱胁迫前的电导率差异不大,都在15%左右;不同嫁接组合处理叶片相对电导率随着干旱胁迫程度的加深都呈上升的趋势,但上升的幅度不同。M/M处理的相对电导率在干旱胁迫处理14 d时最高,而M/1103P相对电导率在此时最低,并且随着胁迫时间的延长其电导率上升幅度最小。从细胞膜透性看,不同砧木品种嫁接马瑟兰的抗旱性表现为M/1103P>M/3309A>M/5BB>M/M。

图1 干旱胁迫对不同嫁接组合叶片相对电导率的影响
2.2 干旱胁迫对不同嫁接组合叶绿素含量的影响
叶绿素是反映植物光合能力强弱的一个重要指标,同时也是参与光合作用过程中的主要物质[13]。不同砧木嫁接苗叶片的叶绿素含量随着干旱胁迫程度的加深均呈现下降的趋势。叶绿素含量在干旱胁迫初期下降幅度小,后期下降幅度大,说明叶绿素的分解随着干旱胁迫的进行加快。
由图2可知,不同嫁接组合处理叶绿素含量在干旱胁迫处理前差异不显著。M/M处理叶绿素在干旱胁迫7 d时出现了明显的下降,而且不同嫁接组合处理间差别幅度很大,其中M/1003P嫁接苗下降的幅度最小。随着干旱胁迫程度的加深,各处理叶片叶绿素含量继续呈现下降的趋势;干旱胁迫第28 d时,M/M嫁接苗叶片叶绿素含量最低,M/1003P嫁接苗叶片叶绿素含量最高。

图2 干旱胁迫对不同嫁接组合叶绿素含量的影响
2.3 干旱胁迫对不同嫁接组合光合特性的影响
由图3可知,不同嫁接组合叶片净光合速率(Pn)随着干旱胁迫程度的加深先升高后降低。各处理的净光合速率在干旱胁迫处理的第7 d时均达到最高;M/1003P和M/3309A嫁接组合在干旱初期Pn上升的幅度较大,说明其抗旱性较强;反之,抗旱性弱的嫁接组合上升的幅度就小,如M/M和M/5BB。到干旱胁迫第21 d时,各嫁接组合的净光合速率已降至最低。

图3 干旱胁迫对不同嫁接组合叶片净光合速率Pn的影响
由图4可知,不同嫁接组合叶片蒸腾速率Tr随着干旱胁迫时间的推移整体呈现下降趋势。马瑟兰自嫁接处理在干旱胁迫前的蒸腾速率较小,但随着干旱胁迫程度的加深,蒸腾速率下降的幅度是最大的,到第14 d时下降速度变缓且幅度变小。M/1003P和M/3309A的蒸腾速率在干旱胁迫过程中下降速度和幅度一直较缓慢。

图4 干旱胁迫对不同嫁接组合叶片蒸腾速率Tr的影响
由图5可知,随着干旱胁迫程度的增加,马瑟兰葡萄不同嫁接组合处理叶片气孔导度先升高后降低。干旱胁迫前叶片气孔导度较高的处理是M/1003P和M/3309A;到干旱胁迫第7 d时,M/1003P、M/3309A及M/M嫁接组合处理叶片气孔导度都呈缓慢上升趋势,但M/5BB处理的气孔导度呈缓慢下降趋势。从干旱胁迫第7 d到第14 d这一周内,各处理叶片气孔导度下降的幅度较大、速度较快;到干旱胁迫第21 d时,各处理叶片气孔导度下降至最低值。
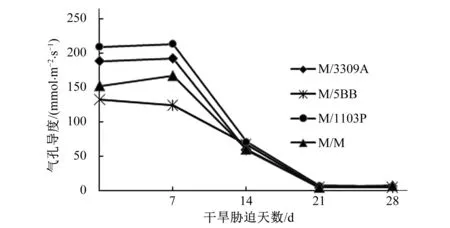
图5 干旱胁迫对不同嫁接组合叶片气孔导度Gs的影响
由图6可知,不同嫁接组合叶片胞间CO2浓度都是随着干旱胁迫程度的增加Ci先降低后升高。叶片的Ci在干旱胁迫初期随着干旱胁迫的推移逐渐降低,但降低的幅度不一样,M/3309A降低幅度最小;到第14 d时,由于干旱胁迫气孔关闭,导致Ci降至最低;在之后的14 d,随着干旱进一步加重,叶片中叶绿体同化CO2的能力下降,导致Ci开始上升。

图6 干旱胁迫对不同嫁接组合叶片胞间CO2浓度Ci的影响
2.4 干旱胁迫对不同嫁接组合旱害指数的影响
植株旱害指数能够反映植株的抗旱性差异。由图7可知,干旱胁迫处理28 d后,马瑟兰不同嫁接组合处理中,M/1103P处理嫁接苗旱害指数最低,与M/3309A无显著性差异,与M/5BB和M/M有显著性差异,而对照各处理无显著性差异,说明M/1103P和M/3309A抗旱性较强。

图7 干旱胁迫对不同嫁接旱害指数的影响
2.5 干旱胁迫对不同嫁接组合株高和径粗的影响
由图8可知,马瑟兰不同嫁接组合干旱胁迫前后都是以M/1103P的株高最高,且与其他处理都有显著性差异。

图8 干旱胁迫对不同嫁接组合株高的影响
由表1可知,马瑟兰不同嫁接组合接穗径粗在干旱胁迫前与对照有显著性差异,干旱胁迫28 d后,M/5BB接穗径粗最大,且与其他各处理有显著性差异。马瑟兰不同嫁接组合干旱胁迫前和干旱胁迫28 d后都是以M/1103P处理砧木径粗最大,与M/5BB处理无显著性差异,但与M/3309A和M/M有显著性差异。

表1 干旱胁迫对不同嫁接组合接穗径粗和砧木径粗的影响
3 结 语
光合作用是植物积累有机物质的重要生理活动,也是植物最基本的生命活动[14,15]。判断植株生长好坏和抗逆性强弱的标准之一就是光合速率的高低[16,17]。植物光合速率高低的限制因子就是水分,水分胁迫会对果树的营养生长和生殖生长造成不可逆的伤害,是因为水分胁迫是导致植物光合速率下降的重要原因[18],因此,在干旱胁迫下仍能保持高水平的净光合速率(Pn)对葡萄的生长和发育具有重要意义[19]。本试验的结果显示,4种嫁接组合随着干旱胁迫程度的加深其净光合速率(Pn)均呈下降的趋势,这与陈丽[20]等人的研究结果一致,其净光合速率(Pn)的大小表现为M/1103P>M/3309A>M/5BB>M/M,说明砧木嫁接能有效提高葡萄的Pn值。
植物叶片中的叶绿素浓度在一定范围内与植物光合速率呈正相关[13]。葡萄叶片中的叶绿素总量会受到干旱胁迫导致叶绿体色素非常不稳定[21]。本试验结果表明,随着干旱胁迫时间的推移和程度的加深,葡萄叶片的叶绿素含量变化趋势同蒸腾速率(Tr)和气孔导度(Gs)的变化趋势一致,即持续降低,其原因一方面可能是因为干旱胁迫导致叶片中的叶绿素合成渠道受阻,另一方面可能是因为葡萄叶片中的叶绿素分解加快。3种嫁接组合及马瑟兰自嫁接苗的叶绿素含量的高低表现为:M/1103P>M/3309A>M/5BB>M/M,说明砧木嫁接能有效提高葡萄的叶绿素含量,从而提高了葡萄的净光合速率,这与前面的研究结果相一致。
综上所述,在重度水分胁迫下,各嫁接组合植株间的生长量、旱害指数、叶绿素、质膜损伤程度以及光和特性差异显著;马瑟兰葡萄用砧木嫁接后栽培能明显提高其抵御干旱的能力,且以不同的砧木嫁接后抗旱性较强的嫁接组合是M/1103P。
