β1-肾上腺素受体自身抗体通过激活内质网应激诱导心肌细胞凋亡*
2018-11-26侯晓鸿何金玲王晓晖
侯晓鸿, 宁 娜, 李 杨, 何金玲, 燕 子, 原 媛, 王 丽△, 王晓晖△
(山西医科大学基础医学院 1病理教研室, 2生理学系, 3形态学实验室, 山西 太原 030001)
心功能不全是各种病因所致心血管疾病的终末阶段,其病因复杂,死亡率高,已成为发达国家和发展中国家的重大公共卫生问题[1],但发病机制尚未完全阐明。大量研究表明,心功能不全的发生发展过程中存在心肌细胞凋亡[2],并且心肌细胞凋亡对心肌结构和功能的调节起着至关重要的作用,因此抑制心肌细胞凋亡可以有效防止和缓解心功能不全[3]。
近年来研究显示,在40%~60%心功能不全患者血清中均可检测到β1-肾上腺素受体自身抗体(β1-adrenoceptor autoantibody,β1-AA)[4],进一步研究发现,β1-AA可以与β1-肾上腺素受体(β1-adrenoceptor,β1-AR)特异性结合,诱导心肌细胞凋亡[5]。然而,β1-AA如何诱导心肌细胞凋亡目前尚不清楚。
内质网在氧化应激、维持钙稳态和触发细胞凋亡信号等多种细胞过程中发挥重要作用[6]。当内质网稳态因各种因素被打破,就会引起内质网应激。轻度和短暂的内质网应激是细胞的一种自我保护机制,而持续严重的内质网应激,会诱发心肌细胞凋亡[7]。然而,内质网应激在β1-AA诱导的心肌细胞凋亡过程中是否发挥作用目前尚不清楚。本研究将通过在体和离体实验,探讨内质网应激在β1-AA诱导的心肌细胞凋亡中的作用,为阐明β1-AA诱导心肌细胞凋亡的机制提供新的理论支持。
材 料 和 方 法
1 动物和细胞
8周龄雄性Sprague-Dawley (SD)大鼠,体重180~200 g,由山西医科大学动物中心提供,合格证编号为SCXK(晋)2015-001。将大鼠置于恒定环境,室温(22±2) ℃、湿度35%~55%,自由饮食。本研究中涉及动物的所有程序均得到山西医科大学伦理委员会批准,使用过程符合国家实验动物使用规定。H9c2心肌细胞系购买自中国科学院上海细胞库。
2 主要试剂
β1-AR细胞外第二环(the second extracellular loop of β1-AR,β1-AR-ECII)抗原肽段由吉尔生化上海有限公司合成;完全弗氏佐剂(complete Freund’s adjuvant, CFA)、不完全弗氏佐剂(incomplete Freund’s adjuvant, IFA)、2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS)和4-苯基丁酸(4-phenoxybutyric acid,4-PBA)均购自Sigma;DMEM培养基和BCA试剂盒购自Thermo Fisher Scientific;抗葡萄糖调节蛋白78(glucose-regulated protein 78,GRP78)、C/EBP同源蛋白(C/EBP homologous protein, CHOP)和caspase-12抗体均购自Abcam;抗GAPDH抗体及 II 抗购自中杉金桥;0.45 μm PVDF膜购自Whatman;SuperECL Plus超敏发光液购自普利莱;SDS-PAGE凝胶制备试剂盒购自索莱宝;胎牛血清购自Excell;CCK-8试剂盒购置同仁化学。
3 主要方法
3.1主动免疫大鼠 选取8周龄雄性SD大鼠,随机分为2组:免疫(β1-AA)组和溶剂对照(vehicle)组,将β1-AR-ECII抗原肽段溶解稀释,并和完全弗氏佐剂1 ∶1混匀后,采用多点背部皮下注射(0.4 μg/g)即首次免疫;以后每隔2周加强免疫1次,并加入不完全弗氏佐剂1 ∶1乳化抗原,共免疫8周;溶剂对照组用等量Na2CO3溶液替代抗原溶液,实验方案同免疫组。
3.2链霉亲和素-酶联免疫吸附实验(SA-ELISA)检测血清中β1-AA水平 将合成的β1-AR-ECII(10 mg/L)包被空白ELISA板,4 ℃过夜;磷酸盐缓冲液封闭ELISA 板;将阳性对照、空白对照及待测血清稀释后加于ELISA板,加带有生物素标记的 II 抗及带有辣根酶标记的链霉卵白素于ELISA板,以上各步分别在37 ℃水浴1 h;底物显色30 min;最后测定吸光度(A)值。计算P/N判定结果,P/N=(标本A值-空白对照A值)/(阴性对照A值-空白对照A值),P/N≥2.1 定为抗体阳性,P/N≤1.5 为阴性。
3.3亲和层析法提纯β1-AA 采用亲和层析法提纯主动免疫大鼠血清中的β1-AA,应用于离体实验研究。从4 ℃取出层析柱放置室温30 min,将三蒸水注入层析柱内部,冲去保存在柱子中的乙醇;用结合缓冲液冲洗柱子后,抽取含有β1-AA的血清滤过柱子并用洗脱缓冲液冲洗柱子,使β1-AA随同洗脱缓冲液滴到EP管中;用BCA试剂盒进行蛋白定量。
3.4Western blot检测心肌组织中GRP78、CHOP和caspase-12的蛋白表达 提取大鼠心肌组织蛋白,用BCA试剂盒进行蛋白定量。上样并进行SDS-PAGE分离以及用PVDF膜转膜,5%的脱脂奶粉封闭,与Ⅰ抗稀释液进行反应,4 ℃过夜;次日加II抗反应2 h;将膜置于全自动曝光仪内,曝光拍摄条带,ImageJ软件分析灰度值,将GAPDH作为内参照,计算不同蛋白的相对表达量。
3.5TUNEL染色检测心肌组织的凋亡水平 取大鼠心肌组织制备切片,脱蜡和水化;用蛋白酶K通透细胞;制备TUNEL反应混合物并标记反应;DAB显色;苏木素复染,封片,显微镜下拍照,Image-Pro Plus 6.0 图像分析软件进行凋亡细胞计数。心肌细胞凋亡率即凋亡指数(apoptotic index,AI;%)=凋亡心肌细胞核数/心肌细胞核总数×100%。
3.6免疫组化法检测心肌组织中GRP78、CHOP及caspase-12的蛋白原位表达 选取大鼠心肌组织浸入福尔马林固定;随后石蜡包埋,切片,二甲苯脱蜡至水;H2O2避光处理及抗原修复;用血清室温封闭;与Ⅰ抗稀释液反应,4 ℃过夜;次日加相应II抗37 ℃孵育20 min; 显色,复染,脱水,透明,然后封片。在显微镜下观察拍照分析。
3.7细胞培养及处理 用含10%胎牛血清的DMEM培养基加入1%青链霉素混合液用于H9c2细胞的培养,将培养瓶置于含5% CO2、37 ℃及饱和湿度的恒温细胞培养箱中,隔天传代。根据具体情况更换新的培养基,加入1 μmol/L的β1-AA进行处理,分别在12 h、24 h和36 h收集样本。

3.9Annexin V-FITC/PI双染流式细胞术检测心肌细胞的凋亡水平 向6孔板中加入细胞悬液(每孔1 mL),待细胞贴壁后,实验组1 mmol/L 4-PBA预处理2 h,再加1 μmol/L β1-AA处理细胞6 h,收集样本加凋亡试剂,用流式细胞术检测并分析细胞凋亡水平。
4 统计学处理
采用SPSS 17.0软件对数据进行统计分析。结果以均数±标准误(mean±SEM)表示,多组间比较采用单因素方差分析,流式细胞术结果组间两两比较采用SNK-q检验,其它结果组间两两比较采用LSD-t检验,以P<0.05为差异有统计学意义。
结 果
1 β1-AR-ECII主动免疫大鼠模型的建立
SA-ELISA结果显示,随着主动免疫时间的延长,对照组大鼠血清中β1-AA的水平无明显变化,而主动免疫2周后大鼠血清中β1-AA的水平与对照组相比显著增加(P<0.01),持续增加至8周(P<0.01),提示主动免疫大鼠模型建立成功,见图1。
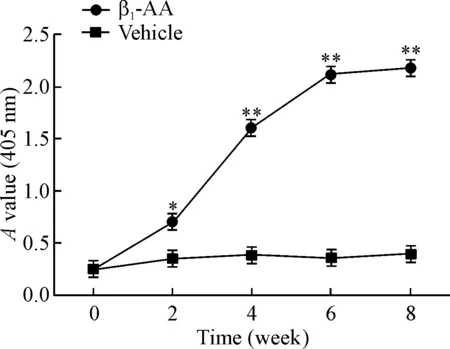
Figure 1.The A value of β1-AA increased in the serum of adult rats in the active immunization model with synthetic peptide. Mean±SEM. n=15. * P<0.05, ** P<0.01 vs vehicle group.
2 β1-AA诱导心肌组织凋亡水平升高
TUNEL染色后,凋亡细胞的细胞核呈现棕黄色,正常细胞的细胞核呈现蓝色。在本研究中,对照组心肌组织可见少量棕黄色细胞核,即存在少量凋亡细胞,而β1-AA主动免疫2周时心肌组织中TUNEL染色阳性即棕黄色的细胞核明显增多,一直持续至主动免疫8周(P<0.01),见图2。以上结果提示β1-AA长期存在可以引起心肌细胞凋亡增加。
3 β1-AA诱导心肌组织内质网应激及其相关凋亡通路激活
Western blot结果显示,与对照组相比,GRP78、CHOP和caspase-12的蛋白表达水平在β1-AA主动免疫4周时开始出现明显增加,β1-AA主动免疫8周时,GRP78、CHOP和caspase-12的蛋白表达水平进一步增加(P<0.01),见图3A。免疫组化结果显示,与对照组相比,心肌组织GRP78、CHOP及caspase-12的蛋白表达在主动免疫4周明显增加,持续增加至8周(P<0.05),与Western blot结果一致,见图3B。以上结果均提示β1-AA长期存在可以诱导内质网应激及其相关凋亡通路激活。
4 β1-AA诱导H9c2心肌细胞活力降低
结果显示,与对照组相比,β1-AA干预H9c2心肌细胞12 h细胞活力开始显著降低,36 h达最低,β1-AA干预48 h心肌细胞活力持续降低(P<0.01),见图4。这提示β1-AA可以引起H9c2心肌细胞活力显著下降。
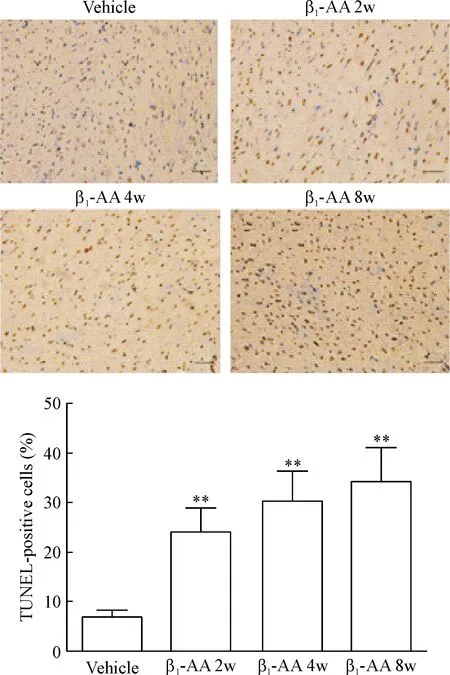
Figure 2.The significant rise in cardiomyocyte apoptosis was induced by β1-AA. The scale bar=40 μm. Mean±SEM. n=6. ** P<0.01 vs vehicle group.
5 β1-AA诱导心肌细胞H9c2凋亡增加
流式细胞术结果显示,与对照组相比,β1-AA干预H9c2心肌细胞6 h细胞的凋亡水平明显增加(P<0.05),见图5。这提示β1-AA可以引起H9c2心肌细胞凋亡水平显著升高。
6 抑制内质网应激可以部分逆转β1-AA诱导的心肌细胞活力降低和凋亡增加
4-PBA预处理心肌细胞2 h可明显逆转β1-AA诱导的心肌细胞活力降低。与β1-AA单独处理组相比,0.5 mmol/L和1 mmol/L 4-PBA处理H9c2 细胞2 h后,再给予β1-AA干预的心肌细胞可见活力明显升高(P<0.05),然而,当2.5 mmol/L和5 mmol/L浓度的4-PBA预处理心肌细胞后,与β1-AA单独处理组相比细胞活力无明显变化,提示4-PBA在0.5 mmol/L和1 mmol/L浓度可以明显改善H9c2细胞活力,其中1 mmol/L 4-PBA对于心肌细胞活力的改善最明显,因此我们选择1 mmol/L 4-PBA 用于后续研究,见图6。Annexin V-FITC/PI双染流式细胞术检测结果发现,与β1-AA单独处理组相比,1 mmol/L 4-PBA预处理H9c2细胞2 h,再给予β1-AA干预,心肌细胞的凋亡水平部分恢复(P<0.05),见图5。
讨 论
本研究中,我们建立β1-AA主动免疫大鼠模型,TUNEL结果显示,主动免疫大鼠心肌组织的凋亡率增加;利用Western blot和免疫组化方法发现β1-AA可以诱导心肌组织内质网应激明显增强;同时采用亲和层析法提纯主动免疫大鼠血清中的β1-AA作用于H9c2细胞,结果发现β1-AA可以显著降低心肌细胞活力,同时凋亡水平明显升高。为进一步探讨β1-AA诱导的内质网应激与凋亡之间的关系,本研究采用4-PBA抑制内质网应激,结果发现抑制内质网应激可以有效降低心肌细胞的凋亡,恢复细胞存活率,提示内质网应激参与了β1-AA诱导的心肌细胞凋亡。
近年来,我国居民心血管疾病的患病率逐渐增加,心功能不全作为心血管疾病的终末阶段,已经成为心血管疾病的主要死亡原因。研究表明,在40%~60%心功能不全患者血清中均可检测到高滴度的β1-AA[4];β1-AA是针对β1-AR-ECII的自身抗体,人鼠同源性高达100%[8]。β1-AA可以与β1-AR结合发挥类激动剂样的作用,引起细胞内cAMP含量积累,进而影响细胞内Ca2+的稳态,引起心肌细胞死亡,最终导致心功能不全[9]。我们课题组前期研究也发现β1-AA长期存在可以导致大鼠心肌重构和心脏功能受损[10];而采用免疫吸附法去除心功能不全患者血清中的β1-AA可以大大改善心功能不全[11]。由此可见,β1-AA可以造成明显的心肌损伤,进而引起心功能不全,然而机制不清。
心肌细胞死亡在心功能不全的发展过程中发挥重要作用。研究报道,心肌细胞持续丢失可引起心功能不全,抑制心肌细胞死亡可有效改善心功能[12]。在本实验中,为了观察β1-AA对H9c2心肌细胞死亡的影响,我们首先建立β1-AR-ECII主动免疫大鼠模型,腹主动脉采血后,采用亲和层析法提纯大鼠血清中的β1-AA,用于后续离体实验研究。本研究发现,1 μmol/L β1-AA作用于H9c2心肌细胞12 h时可以引起细胞活力明显降低,36 h细胞活力降至最低,48 h细胞活力持续降低,提示β1-AA可以诱导心肌细胞死亡。细胞凋亡作为细胞程序性死亡的方式之一,可以维持机体稳态,然而,心肌细胞凋亡的持续存在可以增加心肌细胞死亡,导致心功能不全,采用caspase抑制剂Z-VAD-FMK可以有效抑制心肌细胞的凋亡,进而改善心肌的收缩和舒张功能,提示心肌细胞凋亡在心功能不全中发挥重要作用[13]。β1-AA
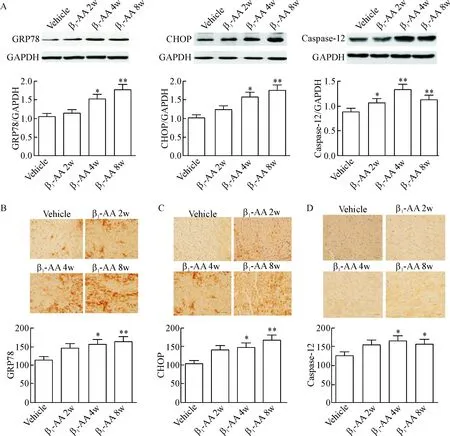
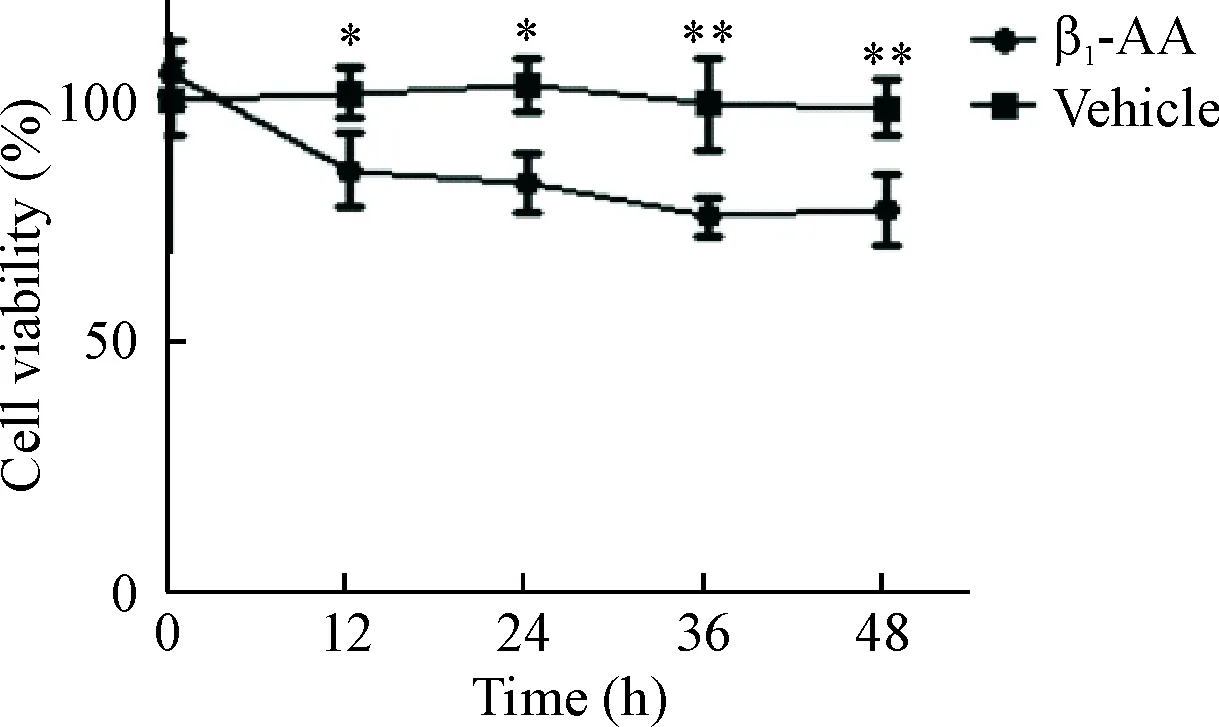
Figure 4.The reduced viability of H9c2 myocardial cells induced by β1-AA. Mean±SEM. n=6. * P<0.05, ** P<0.01 vs vehicle group.
可以激活心肌细胞cAMP依赖的蛋白激酶信号通路,引起caspase-3活化,导致心肌细胞凋亡[14]。本研究发现,β1-AA主动免疫2周时心肌组织中TUNEL染色阳性细胞数明显增多,提示心肌组织凋亡明显增加,同时,β1-AA直接作用于H9c2心肌细胞12 h后凋亡水平显著增加。但是β1-AA如何诱导心肌细胞凋亡目前尚不十分清楚。
目前认为有3条通路诱导凋亡的发生:线粒体通路、死亡受体通路和内质网通路[15-17]。内质网通路是新近发现的途径,持续内质网应激可以诱导内质网-凋亡途径,引起细胞凋亡[18]。内质网对刺激极为敏感,当各种应激诱导其功能紊乱时,诱导内质网应激增强。GRP78是内质网应激特异标志分子[19],CHOP,caspase-12已被证明介导内质网应激-凋亡相关通路的关键分子[20]。因此,本研究选择GRP78、CHOP和caspase-12来反映内质网应激及其相关凋亡通路的变化情况。有研究表明,β1-AR激活可以诱导内质网应激增加,引起内质网标志蛋白GRP78表达增加,同时发现CHOP和caspase-12蛋白表达也增加[21]。而使用β-AR拮抗剂美托洛尔可以有效抑制内质网应激[22]。那么,β1-AA同样作为β1-AR的激动剂,是否也会引起心肌细胞内质网应激激活呢?在本研究中,我们发现,β1-AA主动免疫大鼠2周时心肌组织GRP78、CHOP和caspase-12的蛋白表达与对照组相比无明显变化,而β1-AA主动免疫大鼠4周和8周时GRP78、CHOP和caspase- 12的蛋白表达水平明显升高,提示β1-AA可以诱导内质网应激及其相关凋亡通路的激活。
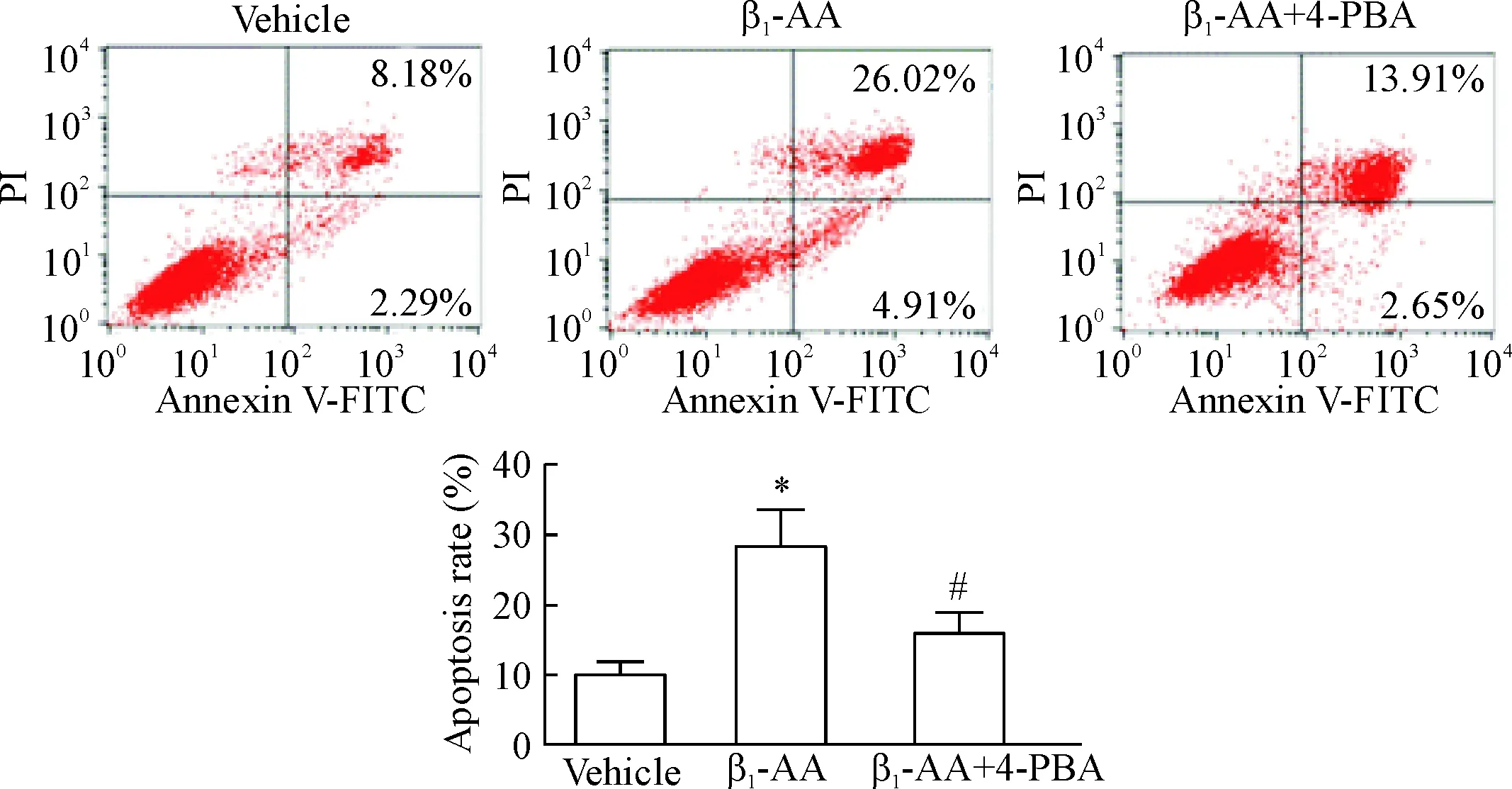
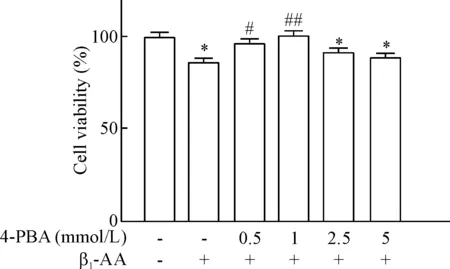
Figure 6.Decreased cell viability induced by β1-AA was partially recovered by endoplasmic reticulum stress inhibitor 4-PBA. The H9c2 myocardial cells was pretreated with 4-PBA (0 mmol/L, 0.5 mmol/L, 1 mmol/L, 2.5 mmol/L and 5 mmol/L) for 2 h before adding β1-AA (1 μmol/L), and CCK-8 assay was performed to measured the cell viability. Mean±SEM. n=9. * P<0.05 vs vehicle group; # P<0.05, ## P<0.01 vs β1-AA group.
内质网应激作为细胞凋亡的主要通路之一,其功能的抑制对细胞凋亡具有改善作用。4-PBA是一种化学分子伴侣,稳定蛋白质构象,改善内质网蛋白质折叠和转运,缓解内质网应激,是内质网应激的特异性抑制剂。有研究表明,4-PBA可以抑制内质网应激改善心肌细胞的凋亡水平[23]。为了进一步验证内质网应激在β1-AA诱导的心肌细胞凋亡中发挥何种作用,本研究采用1 mmol/L 4-PBA预处理H9c2心肌细胞后,再加β1-AA干预,通过Annexin V-FITC/PI双染流式细胞术检测细胞凋亡情况,结果发现4-PBA预处理H9c2心肌细胞后可降低β1-AA诱导细胞的凋亡水平,同时,通过CCK-8法检测细胞的存活率,结果发现4-PBA的预处理可以有效提高β1-AA诱导H9c2心肌细胞的活力。这些结果提示抑制内质网应激可以有效逆转β1-AA诱导的心肌细胞凋亡。
综上所述,本研究证实β1-AA的长期持续存在可以引起心肌细胞活力降低,心肌细胞凋亡水平增加,同时发现β1-AA诱导内质网应激及其相关凋亡途径激活,采用内质网应激抑制剂4-PBA可以有效改善β1-AA诱导的细胞凋亡水平增加和活力下降,提示β1-AA可以通过内质网应激诱导心肌细胞凋亡。因此,早期合理调控内质网应激,进而调控心肌细胞凋亡,可能有效减少心肌细胞丢失,改善心功能。这为治疗β1-AA阳性患者的心功能不全提供新的理论依据。
