氧化应激诱导剂甲萘醌对脱氮假单胞杆菌代谢过程的影响
2018-11-22章帅文李昆太
杨 勇,刘 群,章帅文,李昆太
(江西农业大学生物科学与工程学院,江西南昌330045)
维生素B12(Vitamin B12)又叫钴胺素,属于咕啉类化合物,是唯一含金属的维生素类化合物[1]。作为一种重要的生物活性物质,维生素B12是许多重要生物化学反应所必需的辅酶[2],其参与的生化反应过程包括了DNA的合成和调控、脂肪酸的合成、氨基酸的代谢及能量的产生等[3]。目前,维生素B12主要用于治疗恶性贫血症以及末梢神经炎[4]。
由于维生素B12的分子结构极为复杂,其全部化学合成需要多达70余个的反应步骤[5],且成本昂贵,故目前几乎都是通过微生物发酵的方式来生产维生素B12[6]。自然界中,维生素B12的生物合成存在两种不同的合成途径,即好氧途径与厌氧途径[7]。这两种合成途径的本质区别在于钴离子螯合到咕啉环的时机不同以及中心咕啉环的缩合机制不同[8]。脱氮假单胞杆菌由于具有完整的维生素B12合成基因,可在好氧的条件下完成维生素B12的全合成,因而被广泛开发应用于维生素B12的发酵生产[9]。
假单胞菌属(Pseudomonassp.)就是一类典型的具有ED代谢途径的微生物,如嗜糖假单胞菌(P.Saccharophila)[10],恶臭假单胞菌(P.putida)[11]、荧光假单胞菌(P.fluorescens)[12]等。有研究表明,假单胞菌属对胞内氧化环境较为敏感,其ED代谢途径所产生的NADPH对于增强菌体对抗氧化应激(oxidative stress)反应的能力发挥着重要的作用[13]。例如,Singh等[14]在荧光假单胞菌培养过程中外源添加100 μmol/L的超氧生成剂甲萘醌(menadione),结果表明菌体在氧化应激状态下会显著抑制NAD+激酶(NAD+kinase,NADK)的酶活,并增强 NADP+磷酸酶(NADP+phosphatase,NADPase)的酶活,以增加胞内NADPH 的供给并限制NADH 的合成。
尽管脱氮假单胞杆菌系典型的维生素B12好氧合成菌株,且合成过程中需提供大量的NADPH和ATP参与酶促反应,但其是否会与恶臭假单胞菌、荧光假单胞菌等假单胞菌类一样,对胞内氧化环境较为敏感?其ED代谢途径是否也在抗氧化应激并维持胞内氧化还原平衡方面发挥着重要的调控作用?而甲萘醌作为一种超氧生成剂,可以进入细胞,经单电子还原成半醌自由基,从而进入呼吸链与O2作用形成·O2-[15-16],因此,外源添加甲萘醌可以起到添加活性氧的作用。为此,本文通过考察外源加入不同浓度氧化应激诱导剂——甲萘醌对脱氮假单胞杆菌代谢过程的影响,以期为接下来深入探究脱氮假单胞杆菌在氧化应激状态下的代谢辅因子变化规律,并明确其在维系菌体胞内氧化还原平衡和维生素B12合成中的关键调控作用,为揭示限制性供氧促维生素B12高效合成的代谢机理寻求理论依据。
1 材料与方法
1.1 菌种
脱氮假单胞杆菌(Pseudomonasdenitrificans),由本实验室保存。
1.2 培养基
1.2.1 斜面培养基 蔗糖30 g,蛋白胨10 g,玉米浆10 g,(NH4)2SO40.25 g,(NH4)2HPO41.5 g,MnSO4·H2O 0.1 g,ZnSO4·7H2O 0.1 g,琼脂20 g,去离子水1 000 mL。灭菌前调pH至7.0~7.2。
1.2.2 种子培养基 蔗糖35 g,蛋白胨20 g,KH2PO45 g,(NH4)2SO42.0 g,(NH4)2HPO40.8 g,MgSO41.5 g,ZnSO4·7H2O 0.02 g,MnSO4·H2O 0.2 g,去离子水1 000 mL。灭菌前调pH至7.2~7.4。
1.2.3 发酵培养基 蔗糖50 g,蛋白胨25 g,(NH4)2SO41 g,ZnSO4·7H2O 0.08 g,MgSO42 g,KH2PO40.8 g,甜菜碱10 g,5,6-二甲基苯并咪唑(DMBI) 0.08 g,CoCl2·6H2O 0.15 g,CaCO32 g,去离子水1 000 mL。灭菌前调pH至7.2~7.4。
1.3 摇瓶发酵方法
种子液的制备:每支培养好的脱氮假单胞杆菌新鲜斜面(18×180 mm)加入10 mL无菌水,刮洗制备成菌悬液。以无菌吸管吸取2 mL菌悬液至装有种子培养基的三角瓶(装液量为50 mL/250 mL三角瓶)中,在摇床上进行种子液的培养,培养条件为:温度30 ℃,转速180 r/min,培养至OD700为9~10。
摇瓶发酵:培养好的种子液按10%接种量接种到装有发酵培养基的摇瓶(装液量为40 mL/250 mL三角瓶)中,将无菌过滤好的甲萘醌溶液按照相应的摩尔浓度梯度(浓度梯度分别为0,0.3,0.6,1.0 μmol/L)添加到发酵培养基中,在30 ℃、180 r/min下进行摇瓶发酵培养。
1.4 分析方法
菌体生物量的测定:发酵液取样进行适当稀释,以蒸馏水为对照,于波长700 nm处测定其吸光值。菌体的光密度(OD700)=OD读数×稀释倍数。
总糖测定:苯酚-硫酸比色法[17-18]。
维生素B12含量:取30 mL充分摇匀的发酵液于4 000 r/min离心10 min,去上清后加30 mL蒸馏水搅匀在上述相同条件下离心,倒去上清液后加入10 mL蒸馏水将菌体搅匀,加入质量分数8%亚硝酸钠溶液和冰乙酸各3 mL,摇匀,于95~100 ℃水浴30 min;水浴后冷却至室温,加蒸馏水定容至50 mL,过滤。所得滤液适当稀释后,在波长为361 nm处进行紫外光谱分析,根据所绘制的标准曲线算出维生素B12含量[7]。
1.5 比生长速率,产物比生成速率,基质比消耗速率,产物对基质得率系数
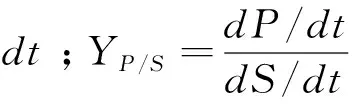
式中:X为菌体生物量浓度(OD值表征);P、S分别为产物浓度与基质浓度,μg/mL;μ为比生长速率,h-1;t表示时间,h。
1.6 试验数据的获得与处理
在发酵培养过程中,每隔24 h取样,分别测定菌体生物量、pH、总糖以及维生素B12含量。每个实验做3次平行实验,取其平均值。
采用Excel 2003进行数据处理,origin 9.0软件进行作图与动力学参数分析。
2 结果与分析
2.1 不同甲萘醌浓度下菌体生长、基质消耗和维生素B12合成的时序性动态变化

图1 甲萘醌对脱氮假单胞杆菌发酵过程基本参数的影响Fig.1 The effects of menadione on basic parameters of Pseudomonas denitrificans fermentation
由图1可以看出,外源添加甲萘醌有利于菌体生物量的形成,其中尤以0.3 μmol/L甲萘醌最佳;在发酵前期,甲萘醌抑制细胞生长可能是因为甲萘醌对于细胞生长以及相关的酶系具有抑制作用,且高浓度甲萘醌抑制作用愈强,而发酵中后期可能菌体开始适应胞外胁迫环境并利用甲萘醌。从pH变化曲线可以看出,添加甲萘醌对发酵液pH初始值基本无影响。在24~48 h高浓度甲萘醌下,pH呈现出先升高后下降趋势,可能是高浓度甲萘醌导致部分菌体凋亡所致。随着发酵的进行,菌体由于生长代谢会持续产生有机酸,导致对照组和试验组pH均出现急剧下降。但是相对于对照组而言,添加甲萘醌能有效延缓发酵液pH的降低,可为菌体生长提供较为适宜的pH环境。随着发酵的进行,菌体代谢不断加剧,发酵液总糖含量呈现明显的下降趋势。与对照组相比,添加甲萘醌加速了菌体的糖耗,再一次反映了添加甲萘醌有利于菌体的生物量形成。另外,随着甲萘醌浓度的增加,维生素B12产量也呈现出明显的降低趋势。尽管0.3 μmol/L甲萘醌浓度下的菌体生物量已达到最大值,但其维生素B12产量仍比对照组降低了7.5%,仅为56.82 μg/mL。
综合以上结果可以得出结论,发酵培养基中添加甲萘醌有利于菌体生长,并可优先延缓发酵液pH的急剧降低,为菌体生长提供适宜的pH环境,但是会显著抑制维生素B12的合成,且甲萘醌浓度越高,抑制作用越强。
2.2 不同甲萘醌添加浓度下的发酵动力学变化
选取0,0.3,0.6 μmol/L这3种甲萘醌浓度下的脱氮假单胞杆菌发酵代谢过程,分别计算出其菌体比生长速率(μ)、总糖比消耗速率(QS)、维生素B12比生成速率(QP)以及维生素B12对总糖的得率系数(YP/S)等动力学参数,结果如表1所示。在整个发酵过程的不同时段,甲萘醌试验组的菌体比生长速率均高于对照组,再次说明了添加甲萘醌会加快菌体生长。其中,添加0.3 μmol/L甲萘醌时,其菌体平均比生长速率明显较高于对照组,说明甲萘醌促菌体生长效果较为显著。由总糖比消耗速率可知,甲萘醌试验组整个发酵过程的总糖比消耗速率要高于对照组,这说明甲萘醌的添加会导致菌体总糖消耗量增加,原因是因为添加甲萘醌情况下,菌体生长代谢加快,因此糖耗加快。在发酵前48 h,由于菌体处于旺盛的对数生长期,总糖比消耗速率明显高于发酵后期。由维生素B12比生成速率综合可以发现,对照组在整个发酵过程的维生素B12平均比生成速率略高于甲萘醌试验组,这说明甲萘醌对产物合成具有一定的抑制作用,可能是因为高氧化应激状态下,脱氮假单胞杆菌不得不将更多的NADPH用于抗氧化应激并维持胞内氧化还原平衡,从而使得流向维生素B12合成途径的NADPH偏少,最终抑制维生素B12合成。此外,分析不同时段维生素B12对总糖的得率系数可知,对照组明显高于试验组,说明甲萘醌试验组的糖耗更多用于菌体生长和代谢维持。
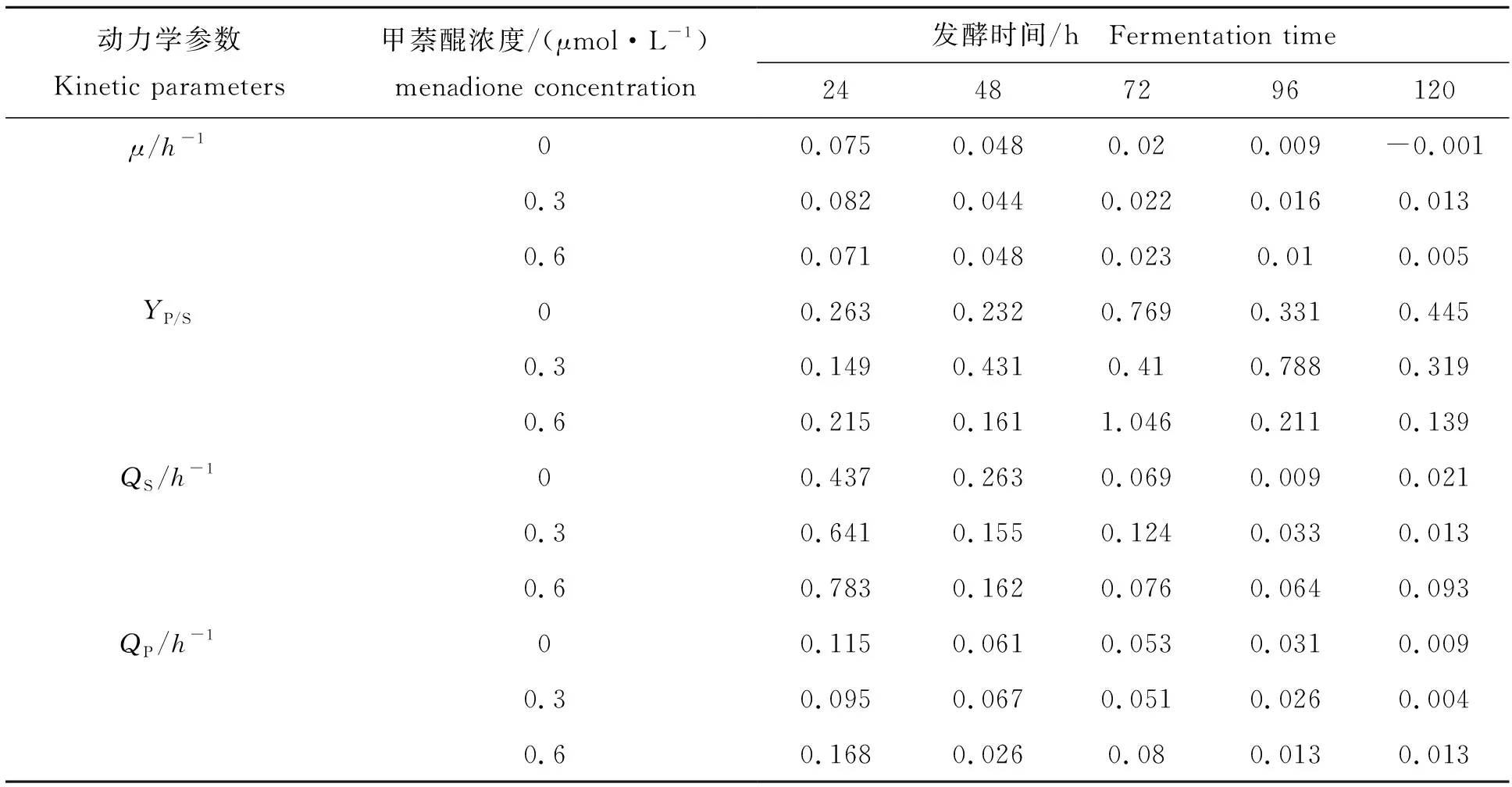
表1 添加甲萘醌对μ、YP/S、QS和QP的影响
3 结论与讨论
微生物通过各种代谢途径提供细胞生长和维持所需的能量、还原力以及不同中间代谢物的需求,其中糖酵解(Embden-Meyerhof-Parnas,EMP)途径是许多微生物的主流糖分解代谢途径,而恩特纳-多多罗夫(Entner-Doudoroff,ED)代谢途径则是某些微生物因缺乏完整的EMP途径而采用的一种替代糖代谢途径[19]。尽管ED途径的产能效率不如EMP途径,但是该途径会生成NADPH,而NADPH恰恰是微生物胞内抗氧化防御系统的重要组成成分,其作为电子供体参与胞内的氧化还原反应以维持胞内正常的氧化还原平衡[20-21]。因此,有些兼性厌氧的环境微生物在进化过程中会选择ED代谢途径分解利用葡萄糖,以对抗并适应其所处的外界氧化环境。
脱氮假单胞杆菌作为典型具有ED代谢途径的微生物,并且Wang等[22]利用13C同位素标记代谢流分析的方法详细考察了维生素B12产生菌脱氮假单胞杆菌的中心代谢网络,发现该菌与其它具有ED代谢途径的恶臭假单胞菌、荧光假单胞菌等假单胞菌类一样,EMP途径的代谢活性很低,主要利用ED途径分解代谢葡萄糖。这似乎表明,脱氮假单胞杆菌对胞内的氧化状态较为敏感。然而,脱氮假单胞杆菌中的维生素B12合成又属于典型的好氧合成途径。那么,胞内氧化状态究竟会对脱氮假单胞杆菌代谢过程,尤其是维生素B12合成过程产生怎样的影响,目前尚不清晰。甲萘醌作为一种强氧化应激诱导剂,王明星等[23]通过研究表明,适量添加甲萘醌能促进过氧化氢酶合成,且当甲萘醌浓度超过5 μmol/L 时,细胞生长明显受到较大程度抑制作用。王业生等[24]研究得出:甲萘醌、二酰胺和H2O2呈浓度和时间依赖性抑制白色假丝酵母(CAL)菌丝形成,且甲萘醌抑制CAL菌丝形成作用最强。
为此,本文通过外源加入不同浓度氧化应激诱导剂甲萘醌,考察了脱氮假单胞杆菌菌体生长、产物合成和基质消耗等发酵过程参数的动力学变化,结果表明:添加甲萘醌可以加快脱氮假单胞杆菌菌体生长、加速总糖消耗,其中以0.3 μmol/L甲萘醌效果较为明显,但是对维生素B12的合成具有一定的抑制作用。结合文献[14]报道,可以分析得出,氧化应激诱导剂甲萘醌的加入虽然能显著增强NADP磷酸酶的酶活,增加胞内NADPH的供给,但同样诱发了脱氮假单胞杆菌胞内氧化环境的加剧,迫使菌体不得不将更多的NADPH用于抗氧化应激并维持胞内的氧化还原平衡,从而使得流向维生素B12合成途径的NADPH偏少,从而最终影响维生素B12的合成量。
