干旱胁迫对草石蚕保护酶活性和渗透调节物质的影响
2018-11-22班甜甜张素勤肖体菊徐彦军耿广东
班甜甜,张素勤,肖体菊,徐彦军,耿广东*
(1.贵州省农业科学院园艺研究所,贵州贵阳550006;2.贵州大学农学院,贵州贵阳550025)
旱害是植物生长过程中常见的危害,它对植物的影响很广泛,可以从种子的萌发期延伸到结果期[1]。干旱首先会使植株叶子出现萎蔫、凋落、黄化等现象,这些现象能直接反应出植物受害程度的大小。根作为植物从外界吸收水分最主要的途径,干旱对其的危害很大[2],如阻碍毛细根的伸长和新生根的形成,降低根系活力,导致植物不能正常生长,甚至停止生长[3]。大量研究表明:水分缺失会导致叶绿素的合成受阻,含量降低[4-5],使叶片上的气孔关闭,阻碍植物对CO2的吸收[6],影响光合效率[7-8]。除此之外在逆境条件下,植物体内脯氨酸含量、可溶性糖含量和可溶性蛋白的含量都会有一定程度的积累以维持细胞的渗透压,帮助植株适应环境。植物的抗氧化酶(SOD、POD、CAT等)保护系统具有清除逆境中产生的O2-、H2O2、OH-等活性氧的功能[9],减轻对细胞膜透性的伤害。MDA作为膜质过氧化最重要的产物之一,对细胞有很大的毒性,MDA积累量常作为衡量植物受害程度的重要指标[10]。除了外部形态的变化,SOD、POD、CAT活性和MDA积累量可以作为评价植物抗性的常用指标[10]。
草石蚕(StachyssieboldiiMiq.)是水苏属多年生草本植物,食用部分为地下茎,营养价值丰富,具有补血、强身、清热解毒、治咳嗽等功效[11]。草石蚕有一定的耐旱能力,但在极端条件下,其产量和品质均会受到很大影响,因此探索草石蚕的抗旱机理,对提高其产量和品质具有重要意义。目前,对植物干旱胁迫条件下的生理生化变化的研究较多,但对草石蚕干旱胁迫方面的相关研究鲜见报道。因此本文以3个抗旱性不同的草石蚕品种为试材,研究干旱胁迫对其保护酶活性和渗透调节物质的影响。本研究拟从生理指标的变化来探讨干旱胁迫对草石蚕的影响,以期了解草石蚕的抗旱机制,并为草石蚕生产中的抗旱问题提供理论指导。
1 材料与方法
1.1 材料
通过前期干旱处理的预试验,由草石蚕的外部形态所受到的影响发现“贵栽1号”、“鲁引1号”和“赣引1号”3个草石蚕品种对干旱的适应能力不同,具体的抗旱能力为:“贵栽1号”(抗旱性强,贵州务川主栽品种)、“鲁引1号”(抗旱性中等,山东滨洲主栽品种)和“赣引1号”(抗旱性弱,江西萍乡主栽品种),本实验以3个抗旱性不同的草石蚕品种为实验材料,研究干旱对草石蚕相关生理方面的影响。
1.2 方法
试验于2016年3月在贵州大学农学院教学实验基地大棚内进行,常规育苗,待幼苗长到5叶1心时,将其移栽到盆中(盆中加等量的土),每盆移栽4棵,每个处理10盆,生长15 d后,进行干旱处理(先浇透水,然后自然干旱),干旱处理4、8、12和16 d后对其进行各种生理指标测定。以正常浇水生长的草石蚕为对照,3次重复。采样时间均为08:00—09:00,试验测定的生理指标有可溶性糖、可溶性蛋白、脯氨酸(Pro)、SOD、POD、CAT和丙二醛(MDA),试验取样材料为从上到下第3和第4片叶。
可溶性糖含量采用蒽酮比色法,可溶性蛋白采用考马斯亮兰 G-250 染色法,脯氨酸测定采用茚三酮比色法,MDA采用硫代巴比妥酸法[12];抗氧化酶SOD、POD和CAT活性的测定采用王学奎方法测定[13]。
1.3 数据处理
采用Microsoft Excel 2013和DPS(V12.01)软件进行数据的处理和分析。
2 结果与分析
2.1 干旱胁迫对草石蚕可溶性糖含量的影响
在干旱条件下,3个品种的草石蚕随胁迫时间延长,可溶性糖含量呈一直上升的变化趋势(图1)。处理第8天时,“鲁引1号”和“赣引1号”可溶性糖含量明显上升,而“贵栽1号”可溶性糖含量上升幅度小,仅比对照提高了5.32%,但与对照达到极显著差异水平;处理第16天时,“鲁引1号”、“赣引1号”和“贵栽1号”可溶性糖含量达到最大值,分别比对照提高了134.07%、80.85%和48.28%,且均极显著高于对照,其中抗旱性强的“贵栽1号”变化幅度最小,抗旱性中等的“鲁引1号”变化幅度最大。

图1 干旱胁迫对草石蚕可溶性糖含量的影响Fig.1 Effect of drought stress on soluble sugar content of Stachys sieboldii Miq.
2.2 干旱胁迫对草石蚕可溶性蛋白含量的影响
由图2可知,随胁迫时间延长,“鲁引1号”和“赣引1号”可溶性蛋白含量呈先升高后降低的变化趋势,而“贵栽1号”则一直升高。干旱处理第4天时,3个品种可溶性蛋白含量变化不明显,与对照未达到显著差异水平;干旱处理第8天时,“鲁引1号”和“赣引1号”比对照有明显的上升,且在第12天时达到峰值,分别比对照上升了78.15%和57.83%,极显著高于对照;处理第16天时较最大值有一定的下降,但仍高于对照。抗旱性强的“贵栽1号”一直升高,第16天时达到最大值,比对照提高了21.23%,差异达极显著性水平。
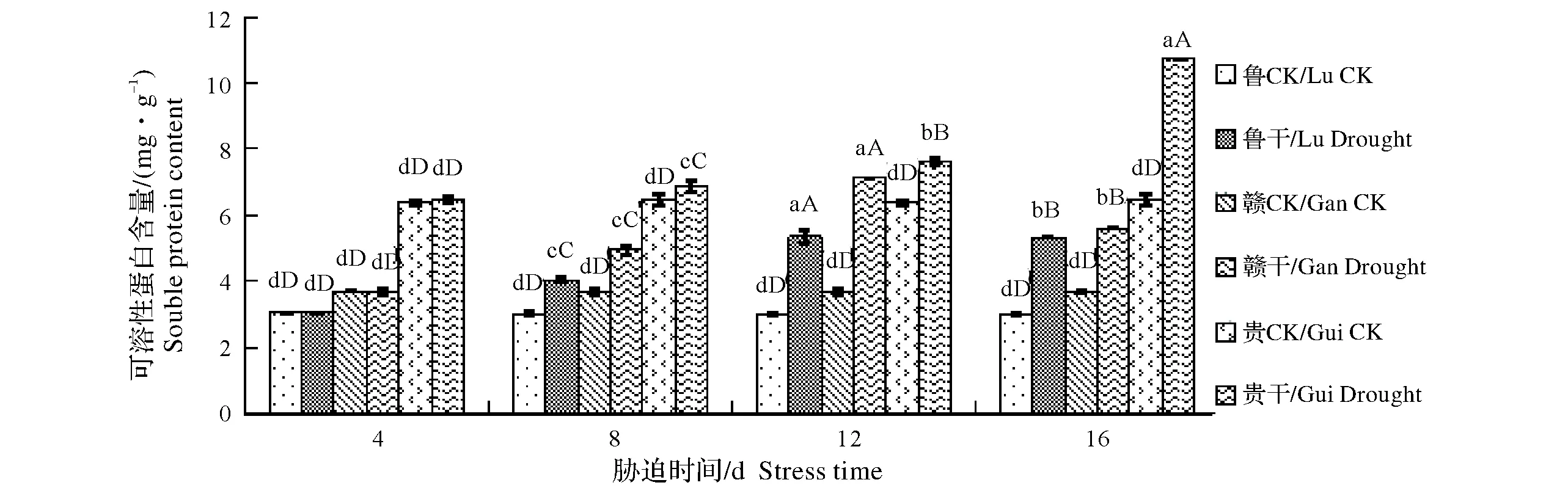
图2 干旱胁迫对草石蚕可溶性蛋白含量的影响Fig.2 Effect of drought stress on soluble protein content of Stachys sieboldii Miq.
2.3 干旱胁迫对草石蚕Pro含量的影响
随干旱胁迫时间延长,3个品种的草石蚕Pro含量呈一直增加的变化趋势(图3)。干旱胁迫第4天时,3个品种的Pro含量升高,且与对照达到极显著性差异水平;干旱胁迫第12天时, Pro含量进一步升高;干旱胁迫第16天时,3个品种的草石蚕Pro含量均达到最大值,与对照相比,“鲁引1号”、“赣引1号”和“贵栽1号”分别增加了164.90%、 203.29%和90.08%。由此可知,草石蚕可通过增加Pro含量来适应旱害,其中抗旱性强的“贵栽1号”Pro含量增加幅度最小,抗旱性弱的“赣引1号”的Pro含量增加幅度最大。

图3 干旱胁迫对草石蚕Pro含量的影响Fig.3 Effect of drought stress on Pro content of Stachys sieboldii Miq.
2.4 干旱胁迫对草石蚕SOD活性的影响
随干旱胁迫时间的延长,3种草石蚕SOD活性均呈一直升高的变化趋势(图4)。干旱胁迫第4天时,SOD的活性变化不明显;干旱胁迫第8天时,3种草石蚕SOD活性均明显升高,“鲁引1号”、“赣引1号”和“贵栽1号”均比对照升高了12.27%、13.63%和13.91%,并均达到极显著性差异水平;干旱胁迫第16 d时,3种草石蚕SOD活性均升高到最大值,与对照相比,“鲁引1号”、“赣引1号” 和“贵栽1号”分别升高了41.67%、35.43%和52.58%,均与第12天处理的达到极显著差异水平。由此可见,抗旱性强的“贵栽1号”升高幅度最大,抗旱性差的“赣引1号”升高幅度最小。

图4 干旱胁迫对草石蚕SOD活性的影响Fig.4 Effect of drought stress on SOD activity of Stachys sieboldii Miq.
2.5 干旱胁迫对草石蚕POD活性的影响
随干旱胁迫时间延长,3种草石蚕的POD活性均呈一直上升的变化趋势(图5)。干旱胁迫第4天时,3种草石蚕POD活性与对照相比升高幅度不明显;干旱胁迫第8天时,3种草石蚕POD活性均有明显的升高,与对照相比均达到极显著差异水平;干旱胁迫第12天时,3种草石蚕POD活性进一步升高,与对照相比,“鲁引1号”、“赣引1号”和“贵栽1号”分别升高了52.21%、82.05%和50.79%;干旱胁迫第16天时,3种草石蚕POD活性均达到最大值,“贵栽1号”的POD活性最高,“鲁引1号”次之,“赣引1号”升高幅度最小。由此可见,抗旱性强的“贵栽1号”POD活性升高幅度最大,抗旱性差的“赣引1号”升高幅度最小。
2.6 干旱胁迫对草石蚕CAT活性的影响
随干旱胁迫时间延长,3种草石蚕的CAT活性均呈一直上升的变化趋势(图6)。干旱胁迫第4天时,3种草石蚕CAT活性与对照相比变化不明显;干旱胁迫第8天时,3种草石蚕CAT活性与对照的变化明显,“贵栽1号”升高了91.90%,“赣引1号”升高了27.89%,“鲁引1号”升高了40.00%,且均达到极显著差异水平;干旱胁迫第16天时,3种草石蚕CAT活性均升到最大值,“鲁引1号”、“赣引1号”和“贵栽1号”分别比对照升高了124.95%、81.37%和205.81%。由此可见,抗旱性强的“贵栽1号”SOD活性升高幅度最大,抗旱性弱的“赣引1号”升高幅度最小。

图5 干旱胁迫对草石蚕POD活性的影响Fig.5 Effect of drought stress on POD activity of Stachys sieboldii Miq.

图6 干旱胁迫对草石蚕CAT活性的影响Fig.6 Effect of drought stress on CAT activity of Stachys sieboldii Miq.
2.7 干旱胁迫对草石蚕MDA积累量的影响
3种草石蚕MDA积累量随干旱胁迫时间的延长呈一直递增的变化趋势(图7)。干旱胁迫第4天时,3种草石蚕MDA积累量与对照相比变化不明显;干旱胁迫第8天时,3种草石蚕MDA积累量较对照增加幅度明显,“赣引1号”、“贵栽1号”和“鲁引1号”分别增加了88.87%、49.76%和112.60%,与对照和第4天的积累量相比均达到显著性差异水平;干旱胁迫第16天时,3个品种的草石蚕MDA积累量达到最大,“赣引1号”、“贵栽1号”和“鲁引1号”分别较对照增加了311.07%、150.71%和269.66%。由此可见,抗旱性强的“贵栽1号”升高幅度最小,抗旱性差的“赣引1号”升高幅度最大。

图7 干旱胁迫对草石蚕MDA积累量的影响Fig.7 Effect of drought stress on MDA content of Stachys sieboldii Miq.
3 讨论
3.1 干旱胁迫对植物渗透调节物质的影响
渗透调节是植物适应干旱胁迫的主要生理机制,渗透调节最初是通过降低细胞内部束缚水的自由能,从而尽量维持细胞内外的水势差,保证细胞在外界水势很低的条件下继续吸水,保证植物体内水的平衡,使植物在干旱环境中能尽量正常的生长,弥补环境缺水对植物造成的影响,对干旱胁迫下植物的生长是非常有益的。干旱时植物呈缺水状态,所以在干旱胁迫下植物会通过积累渗透调节物质来提高细胞浓度[14],降低细胞的渗透势,增强植物从外界吸收水分的能力,以适应干旱的环境[15-17]。抗旱能力不同的植物在干旱环境中积累渗透调节物质的能力不同,所以通过研究渗透调节物质的积累量可以从生理层面说明干旱对不同抗旱能力植物的影响,并解释植物的抗旱能力与渗透调节物质积累量之间的联系。植物细胞的渗透调节物质主要包括Pro、可溶性糖和可溶性蛋白等。本研究测定了干旱胁迫条件下草石蚕叶片内可溶性糖、蛋白、脯氨酸含量及其变化,拟具体解释干旱对不同抗旱性植株的影响。
本试验发现,随干旱胁迫时间的延长,3种草石蚕可溶性糖含量一直升高,“赣引1号”的可溶性糖含量增加最多,“贵栽1号”增加最少。前期可溶性糖含量升高可能是因为干旱胁迫下植物光合系统的运转机能受到影响,植物体内大分子化合物的合成受到抑制,转而合成低分子量的蔗糖等化合物[18];而干旱胁迫后期,植物叶片的含水量大幅度降低,致使可溶性糖的相对浓度升高从而含量升高[19],也可能是因为植物内部机能遭到破坏,生命活动降低,导致对糖的消耗降低,最终使可溶性糖含量升高。本实验中抗旱性强的“贵栽1号”可溶性糖含量增加最少的原因可能是干旱对其影响不大,此时的干旱程度对“贵栽1号”来说并不严重,只需通过积累相对较少的可溶性糖含量就能保证其在干旱逆境中生长,也可以说是因为“贵栽1号”叶片水分的丧失小于其他品种,所以其对可溶性糖积累的需要并不迫切,所以其对干旱的反应表现不如其他品种明显。
干旱胁迫时Pro的合成和积累会急剧增多,有时甚至会增加几十倍,Pro的疏水端具有与蛋白质结合的能力,亲水端又可以和水结合,因此蛋白质就可以利用Pro来束缚大量的水,防止胁迫条件下蛋白质发生脱水现象,造成其性质改变[17],所以Pro对植物的生长有积极作用。本研究中3个品种的草石蚕随着干旱胁迫时间延长,Pro积累量一直上升,前期是因为草石蚕积极调节内部渗透物质,以适宜外部干旱逆境,但随干旱胁迫时间延长,Pro含量的增加很大程度上是因为叶片失水造成的。3个品种脯氨酸含量的变化趋势与可溶性糖含量的变化一致,变化幅度与抗旱能力呈负相关。研究表明在干旱环境中,脯氨酸含量的变化与抗旱性的关系比较复杂,很多研究结果并不一致,所以本文3种草石蚕干旱逆境中脯氨酸含量的变化并不代表其他物种有相同的变化,植物的抗旱能力与很多因素有关,并不能单凭这一指标说明问题。
植物中的蛋白质是重要的有机物之一,主要以各种酶和贮藏蛋白的形式存在于植物体内,在干旱胁迫下,植物的内部调节系统会启动与逆境有关的一些基因,从而合成胁迫蛋白来适应逆境。本实验中随干旱胁迫时间延长,3个品种草石蚕可溶性蛋白含量均有所增加,与张明生等[20]对甘薯研究得到的结论一致,是植物对逆境做出的一种适应性反应,干旱胁迫时间达到一定程度时开始下降,这可能是由2个原因造成的,一是蛋白质的合成能力降低[21],二是分解速度加快[22]。抗旱性强的“贵栽1号”在试验胁迫范围内,随干旱胁迫时间延长,可溶性蛋白含量一直上升,说明其自我调节能力好,在试验中的干旱逆境并没有达到他的干旱适应性临界点,也就是说“贵栽1号”可以适应更加严峻的干旱环境,它对干旱的忍受能力更强。
3种草石蚕的不同渗透调节性物质的变化趋势并不完全一致,但这并不矛盾,因为生物体内的代谢反应是一个复杂的系统,一种物质的变化受很多代谢反应共同影响的结果,所以植物抗旱能力是植物体内各种物质共同作用的结果,并不能以一种物质的变化为标准来判断。
3.2 干旱胁迫对植物抗氧化酶活性的影响
干旱胁迫时植物会产生伤害植物细胞的活性氧,但是植物体内具有清除活性氧的防御保护体系,植物体内的活性氧自由基清除剂抗氧化酶体系(SOD、POD、CAT),在逆境中能使植物通过提高其活性,来减少活性氧对植株的伤害,维持细胞膜的稳定性。干旱胁迫下POD活性升高可以保护植物细胞膜的结构,其活性升高的原因是活性氧的增加刺激了清除酶系统的活性[23]。SOD可以把植物体内有害的超氧自由基转化为过氧化物,再由POD和CAT转化为无危害的水[24]。CAT可以清除细胞内的有毒物质,使细胞免受毒害[25],所以3种酶的活性是鉴定植物抗逆性的重要指标,酶活性越大清除危害物的能力就越大。本试验不同草石蚕品种SOD、POD和CAT活性随干旱胁迫时间的延长一直升高,说明在试验胁迫范围内,草石蚕一直积极调动抗氧化酶活性降低细胞内有毒物质的积累,以适应外界的干旱胁迫,但其调节能力是有限的,当达到一定胁迫程度后,其活性会降低,失去调节能力,植物会受到严重伤害,从本试验结果看,试验范围内的胁迫还未达到这个程度。抗氧化酶的活性对植物对逆境的适应有积极的影响,一般来说抗逆性好的植物的抗氧化酶活性比抗逆性差的植物的抗氧化酶活性要高,本实验结果与此一致,抗旱能力强的“贵栽1号”的抗氧化酶活性高于另外2个品种。
3.3 干旱胁迫对植物体内MDA含量的影响
MDA是植物细胞内膜脂过氧化的产物,会引起细胞膜功能紊乱[26],因此,MDA的积累量可作为评价植物在逆境下受伤害程度的重要指标。MDA的积累量越少,植株受到的危害就越轻,抗性越好,MDA 的积累量越多,植物组织受到的伤害就越严重,抗性越差。本试验中,3个草石蚕品种的MDA积累量变化情况不同,但均成一直升高的变化趋势,这与在花生[27]、白桦[28]和红松[29]等植物上的研究结果相似。抗旱性强的“贵栽1号”MDA积累量最少,所受伤害最小,而抗旱性差的“赣引1号”MDA积累量最多,植株受到的伤害最大。
文中3个草石蚕品种抗氧化酶活性的变化与MDA积累量的变化成负相关,结果与理论相符,正是由于高的酶活具有高的清除氧化产物的能力,才导致抗旱能力强的“贵栽1号”MDA积累量最低。
本文研究了3种不同抗旱性的草石蚕品种在干旱胁迫下,各生理指标的变化,生理的最终作用表现在形态上,但是生理的变化是由分子层面的基因表达决定的,所以后期还可以做相关基因的荧光定量表达,进一步的解释干旱对植株的影响。本文理论上从生理角度解释了抗旱性不同的3种草石蚕在干旱环境下生理上的变化,从实际上为有关3种草石蚕的引种提供了参考依据。
