好氧反硝化技术处理水产养殖废水研究进展
2018-11-19黄志涛江玉立宋协法
黄志涛, 江玉立, 宋协法
(中国海洋大学水产学院,山东 青岛 266003)
水产养殖过程中部分饲料在鱼类消化、吸收等代谢过程中转化为氨氮排泄到养殖环境,产生的残饲、粪便也会在微生物作用下矿化为氨,进而转化成亚硝酸盐氮、硝酸盐氮。循环水养殖过程中的生物过滤环节可以将水体中毒性较高的氨氮和亚硝酸盐氮转化成毒性较低的硝酸盐氮,引起水体中的硝酸盐积累,其质量浓度可达100 mg/L以上,甚至是500 mg/L以上[1]。研究表明,硝酸盐氮对于养殖对象的毒害作用比同质量浓度的氨氮和亚硝酸盐氮要低,但高质量浓度的硝酸盐氮仍会导致鱼类生长缓慢,存活率降低,免疫力下降[2-6]。含有高质量浓度硝酸盐氮养殖废水的排放,是自然环境水体富营养化的重要原因之一,因而应将循环水养殖系统中硝酸盐氮的质量浓度控制在合理范围内。
生物脱氮技术是目前处理水产养殖废水的有效方式之一。实际生产中,养殖水体中的溶氧一般保持在4~8 mg/L。而传统反硝化理论认为氧气的存在会抑制反硝化酶活性,只有在缺氧或兼性厌氧的条件下反硝化酶才具有活性,生物反硝化过程才能进行。20世纪80年代,Robertson等[7]报道了好氧反硝化细菌和好氧反硝化酶系的存在,为水产养殖在有氧条件下去除硝酸盐提供了一种崭新的思路。近年来,随着对生物脱氮研究的深入,好氧反硝化细菌的研究日益增多,但在水产养殖废水处理方面应用研究较少。
本文介绍了好氧反硝化细菌的作用机理,分析了好氧反硝化细菌的影响因素,重点介绍了好氧反硝化细菌在水产养殖废水处理中的应用现状,指出存在的问题并对其前景进行展望,以期为水产养殖废水反硝化的应用和推广提供理论参考。
1 好氧反硝化作用机理探讨
近年来,不同学者对于好氧反硝化作用机理存在不同的观点,本文从微环境、生物化学和微生物3个方面对近几年国内外学者所做的一些研究成果进行阐述。
1.1 微环境理论
微环境理论主要侧重于从物理角度阐述好氧反硝化作用机理[8-9],认为由于氧气的扩散作用,微生物絮体内部或者生物膜表面会产生溶氧的质量浓度梯度,即微生物絮体表面溶氧的质量浓度较高,好氧硝化细菌发生反应,当深入絮体内部时,氧气的传递受到阻碍,其微环境为缺氧区,厌氧反硝化反应占据优势地位。当微生物反应池内溶氧质量浓度不高时,会使缺氧环境的比例有所升高,有利于反硝化作用的进行[10]。实际生产中,由于外部环境多变,微生物群落复杂以及电子传递不均匀等因素,使得在微生物絮体内或者生物膜表面产生多种微环境。
1.2 生物化学机理
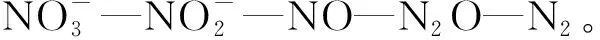
1.3 微生物理论
1.3.1 协同呼吸理论
协同呼吸理论[13]认为,在好氧反硝化过程中,硝酸盐和氧气均可作为电子受体参与反应,细胞色素c和细胞色素aa3之间电子传输过程中的“瓶颈”可被克服,因而电子流就可以同时传输给反硝化酶和氧气,故反硝化就可以在好氧环境中发生(图1)。Huang等[14]也认为,氧气和硝酸盐氮同时存在时,反硝化作为辅助电子传递途径,是对有氧呼吸的补充,可防止烟酰胺腺嘌呤二核苷酸磷酸(NADP)的大量积累。此外,细胞色素的氧化还原水平会控制电子流向不同的细胞色素,以确定电子是否能够发生反硝化。Wilson等[15]提出的好氧反硝化过程中电子传递见图1,反硝化细菌可以将电子从被还原的物质传递给O2,同时也可通过硝酸盐还原酶将电子传递给硝酸根。
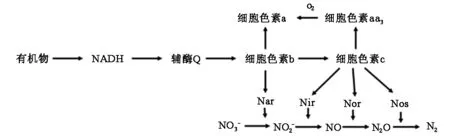
注:Nar为硝酸盐还原酶;Nir为亚硝酸盐还原酶;Nor为一氧化氮还原酶;Nos为一氧化二氮还原酶
1.3.2 好氧反硝化酶作用
好氧反硝化被认为是一个涉及4种酶,即硝酸盐还原酶(Nar)、亚硝酸盐还原酶(Nir)、一氧化氮还原酶(Nor)和一氧化二氮还原酶(Nos)的4步复杂生化还原反应。从好氧反硝化酶系角度阐述好氧反硝化机理开始于对泛养硫球Thiosphaerapantotropha[16](现更名为脱氮副球菌Paracoccusdenitrifications)的研究,在T.pantotropha内包含着膜内硝酸盐还原酶(Nar)和周质硝酸盐还原酶(Nap),在好氧条件下,Nap优先被还原,而在厌氧条件下,Nap的活性被抑制,Nar活跃,被优先利用。Bell等[17]利用Nar对含氮化合物在不同条件下敏感性进行试验,发现在厌氧环境下,90%的Nar活性被抑制,而在好氧的环境下,好氧生长细胞被抑制了25%。Nir主要有2种,一种是人们熟知的细胞色素cd1,由Nirs基因编码;另一种是Nir,位于外周胞质,是一种可溶性含铜酶,由Nirk基因编码。刘兴等[18]研究铜绿假单胞菌YY24发现,该菌存在膜结合Nar,Nir类型为细胞色素cd1型Nir。Nos已在P.stutzeri、P.denitrifications等细菌中分离提纯,其部分特征已被确定由Nor CB基因编码[19]。好氧反硝化菌的Nos是一种含铜蛋白,位于膜外周质中[20]。Bell等[17]认为,在氧气存在条件下脱氮副球菌细胞Nos具有活性,且能将NO、N2O两种气体同时还原。硝酸盐降解过程是因为好氧反硝化菌菌体内依次产生Nar,Nir,Nor和Nos,最终生成氮气。
2 好氧反硝化影响因素
影响好氧反硝化反应的因素有碳氮比(C/N)、溶氧(DO)、碳源、温度、pH等,但不同的菌株种类、好氧反硝化反应器结构以及环境调控措施使得它们的影响能力各不相同。近几年的研究表明,在水产养殖废水处理中,反硝化主要受碳源、溶氧和碳氮比等因素的影响[12,14,21]。
2.1 碳源
碳源是细菌生长过程中必不可少的能量来源,碳源越充足,好氧反硝化速率越快,总氮去除率也越高[21]。孙庆花等[22]从海洋沉积物中分离出一株好氧反硝化细菌,即克雷伯氏菌Klebsiellasp.,发现以柠檬酸三钠为碳源时,硝酸盐氮的去除率为90.27%,且无亚硝酸盐氮积累。Hamlin等[23]利用甲醇、乙酸、葡萄糖、水解的淀粉作为反硝化的碳源对西伯利亚鲟鱼养殖水体进行净化试验,发现4种碳源均能有效地将硝酸盐氮质量浓度从11~57 mg/L降至接近0的质量浓度,在不考虑碳源限制的条件下,每天最大的反硝化率为670~680 g/m3。白洁等[24]从胶州湾沉积物中分离出1 株异养硝化—好氧反硝化菌株Zobellellasp B307,在最优条件下考察其在单一和混合氮源中的脱氮效果,发现最佳碳源为丁二酸钠,在最佳条件下对氨氮和硝酸盐氮的去除率分别达到97.67%和94.39%。Duan等[25]研究发现好氧反硝化细菌VibriodiabolicusSF16的最佳碳源为乙酸钠。Bernat等[26]利用活性污泥处理无碳源废水,脱氮率为1.54 mg/L,而处理投加醋酸钠的废水,脱氮率达到22.50 mg/L,说明好氧反硝化细菌外源呼吸比内源呼吸效率高。可见好氧反硝化细菌最适宜的碳源不尽相同,这是由于碳源结构及分子量的差异导致菌株生长和脱氮效果的不同。
2.2 溶氧
溶氧通过抑制反硝化酶的合成、活性或者与硝酸盐竞争电子供体等途径影响好氧反硝化反应进程,对细菌的反硝化性能至关重要[27]。邵晴等[28]分离的好养反硝化细菌A1,在溶氧为5 mg/L时,对亚硝酸盐的去除率高达99%,且反硝化主要发生在菌种生长的对数期。Huang等[14]认为溶氧是细菌进行好氧反硝化的关键因素,当溶氧为5 mg/L时,好氧反硝化细菌Citrobacterdiversus的反硝化速率最高,溶氧的升高和降低都会造成好氧反硝化速率的下降,原因可能是溶氧在阈值以上时氧气对于反硝化酶有抑制作用。溶氧阈值[29]是区分厌氧反硝化与好氧反硝化的主要标志之一,该值越高,表明菌株对氧的耐受能力越强,即在高溶氧条件下仍表现出良好的反硝化能力。
2.3 碳氮比
水产养殖废水通常是贫营养环境,所以需要额外添加碳源,因而碳氮比(C/N)在反硝化过程中起着重要作用。好氧反硝化的效率随C/N的增加而提高,C/N的降低会减少反硝化基因表达,没有足够的碳源来合成反硝化酶,影响菌体生长以及硝酸盐氮和亚硝酸盐氮的去除[29]。Guo等[30]对好养反硝化细菌HNR进行正交实验表明,在C/N为13时具有最大反硝化率。降低碳氮比,会使反硝化反应过程不完全,导致水体中硝酸盐氮的积累[31]。Van等[32]以乙酸为反硝化碳源实验时发现,当C/N为2∶1或3∶1时,亚硝酸盐氮完全还原成气体,而当C/N下降为1∶1时,仅有50%硝酸盐氮被还原。除此之外,也有研究发现,某些特定的菌株,其C/N的变化范围较小,在最佳C/N值时能够诱导菌株释放足够的反硝化酶,继续增加碳源,其反硝化活性不会增加[33]。梁书城[34]通过改变碳源质量浓度调整C/N分别为5、10、15、20,发现在碳源充足的前提下,DL-23菌株的反硝化能力在C/N 10到15之间均表现出了相似的脱氮效果,继续增加碳源,硝酸盐氮去除能力并未增强。
3 好氧反硝化反应器处理养殖废水应用
循环水养殖系统作为近年来流行的养殖方式,其系统内硝酸盐的积累可达400~500 mg/L[38],因此许多研究者为去除循环水养殖系统中的硝酸盐氮而建立好氧反硝化反应器。Kaiser等[39]在封闭循环式鱼类养殖虹鳟系统中建立一种转盘式生物过滤器,通过添加可水解玉米纤维碳源来促进生物滤池内好氧反硝化细菌进行反硝化,运行118 d,硝酸盐氮由120 mg/L下降至10 mg/L后趋于稳定。Yossi等[40]在金头鲷(Sparusaurata)循环水养殖系统中利用海藻酸盐凝胶微球建立移动床反应器(MBB),以淀粉为碳源处理养殖排放污水,最大硝酸盐氮脱氮速率可达2.7 mg/(h·L)。
4 固定化好氧反硝化细菌处理水产养殖废水

许多学者还将好氧反硝化细菌简单组合构建成微生物制剂,相对于纯菌的处理工艺,微生物制剂的混菌降解能力增强。陈进斌等[44]从天津某海水养殖场底泥中分离筛选3株具有高效好氧反硝化特性的微生物,通过正交试验确定三株菌的最适比例是3∶2∶1,最适比例的菌剂组合在12 h内对海水养殖废水中硝酸盐氮、亚硝酸盐氮的去除率分别为60.45%和67.44%。杜刚等[45]利用PVA和SA固定由光合细菌、放线菌、硝化菌、好氧反硝化菌以及枯草芽孢杆菌组合成的混合菌群用于养殖污水脱氮处理,结果表明氨氮、亚硝酸盐氮、硝酸盐氮的去除率分别为85.0%,87.3%,79.3%。不同功能的微生物固定在同一个载体上,能够构建成一个微型人工生态系统,菌种之间协同作用进行水产养殖废水脱氮,具有良好的去除效果。因此,在水产养殖废水处理中可以将反硝化细菌和硝化细菌按一定比例混合,从而加快脱氮处理效果,实现在同一个反应器中同步硝化反硝化。
5 外加碳源的好氧反硝化处理水产养殖废水
近年来,许多学者利用新型的可生物降解聚合物作为反硝化碳源和生物膜载体去除养殖废水中硝酸盐氮。Boley[48]等利用聚己内酯(PCL)、聚-β-羟丁酸(PHB)和二羧酸二元醇(Bionolle)这3种可生物降解材料作为生物膜载体和反硝化碳源,出水硝酸盐氮质量浓度稳定在0.1 mg/L以下,而亚硝酸盐氮质量浓度也可以维持在0.05 mg/L以下,并未出现积累。张兰河等[49]利用聚羟基丁酸戊酸共聚酯(PHBV)作为固体碳源和生物膜载体,其连接的固相反硝化反应器能显著降低循环水系统中积累的硝酸盐,并维持在较低水平(小于10 mg/L)。
6 结论与展望
好氧反硝化作为一种新型的脱氮工艺,具有技术简单、占地小、能够实现硝化与反硝化同时进行等优点,被国内外学者广泛关注。目前越来越多的好氧反硝化细菌被发现,但对于好氧反硝化技术的研究仍停留在试验阶段,虽然有不少学者将好氧反硝化菌应用到反应器中,但实际应用较少。今后的研究重点应该在以下几个方面:1)筛选分离耐高氨氮、耐高溶氧、耐盐等特性的好氧反硝化细菌,探究影响好氧反硝化细菌脱氮性能的因素,并结合水产养殖废水的特点,设计适合水产养殖的反应器和运行模式,为其在海水循环水养殖系统中的应用打下良好的基础。2)利用分子生物学等技术,深入研究好氧反硝化反应机理,建立好氧反硝化动力学模型,利用基因重组等分子生物学技术强化功能基因,对好氧反硝化细菌种群进行鉴定和跟踪,提高对水产养殖废水的脱氮效率。3)生物固定化技术可以提高水产养殖废水中菌种的密度,防止流速过快、菌种流失,阻碍养殖废水中的有毒物质对菌株的毒害,故应对固定化微生物技术进行深入研究,并将其应用于水产养殖实践中,高效、经济地实现养殖废水脱氮。
□
