洞庭湖水稻土有机氮组分及其与可矿化氮的关系特征
2018-11-05李文军杨奇勇彭保发
李文军,杨奇勇,赵 迪,彭保发
(1.湖南文理学院洞庭湖生态经济区建设与发展湖南省协同创新中心,湖南 常德 4150002.中国科学院南京土壤研究所,土壤与农业可持续发展国家重点实验室,江苏 南京 210008;3.中国地质科学院岩溶地质研究所,广西 桂林 541004)
氮是作物最重要的营养元素,是农田生态系统初级生产力的关键限制养分[1]。土壤有机氮矿化是作物氮素营养供应的重要环节,在未施肥条件下,除起始矿质氮外,作物可利用的土壤氮素主要是可矿化氮[2]。可矿化氮反映土壤氮素的生物有效性[3],其供应特性受土壤有机氮组分结构深刻影响[4]。因此,对土壤有机氮进行分组并分析其组分与可矿化氮间的关系是深入理解土壤氮素矿化本性及有效性的重要基础[5-6]。热酸解程序是经典的土壤有机氮分组技术,其将土壤有机氮划分为酸解氨基酸氮、氨态氮、氨基糖氮、未知氮和酸未解氮5种组分[7]。一系列研究显示土壤酸解氮与土壤可矿化氮关系密切,然而土壤酸解氮中不同组分对土壤矿化氮的贡献效应却存在较大不确定性:不同类型土壤有机氮组成及形态转化特征的差异使得酸解氮各组分甚至酸未解氮都可能单独或共同对土壤可矿化氮有重要贡献[8-11],即使同一类土壤在不同培养条件下发生矿化的主要有机氮组分亦有明显差别[12]。当前大多研究注重利用相关分析来说明土壤有机氮组分与可矿化氮间的关系[4,9,11],然而考虑到土壤有机氮形态转化的复杂性,该法并不能完全揭示它们之间的真正关系,而结合相关分析、多元回归分析和通径分析可更明确地解析土壤有机氮组分对可矿化氮的贡献,但这方面的研究尚少,且主要应用于旱地土壤[7,12]。洞庭湖区是我国重要的双季稻生产区,水稻土是湖区最重要的土壤类型,其面积约占湖区耕地总面积的80%[13],当前对该区水稻土有机氮组成及其生物有效性特征尚不清楚。因此,本研究采集洞庭湖区不同发育类型水稻土,分析其有机氮组成特征并综合应用相关分析、多元回归分析和通径分析研究区域水稻土有机氮组分对可矿化氮的贡献特性,为施氮决策及深入理解区域土壤有机氮素生物有效性提供理论依据。
1 材料与方法
1.1 供试土壤
2012年晚稻收获后在环洞庭湖区(111°13′~113°31′E,28°12′~29°32′N)采集30个表层(0~20 cm)水稻土样本(图1),所有采样点均长期实行早稻-晚稻-冬闲的耕作制度及类似的田间管理措施。所采土壤代表了洞庭湖区由河湖沉积物、第四纪红粘土、花岗岩、石灰岩、紫色砂岩等多种成土母质及冲积平原和丘陵坡岗两种典型地形影响发育形成的水稻土,包括淹育性、潜育性、潴育性3个水稻土亚类及20余个水稻土土种。各水稻土亚类均采集10个土壤样本,采集时,在每个采样点0.1 hm2代表性范围内随机多点采集5 kg土样组成混合样品。采集的混合土样去除肉眼可见石粒和植物残渣,经风干、过2 mm筛且充分混匀后常温下贮存待测。土壤采样点及基本理化性质见表1。
图1 研究区位置与采样点分布
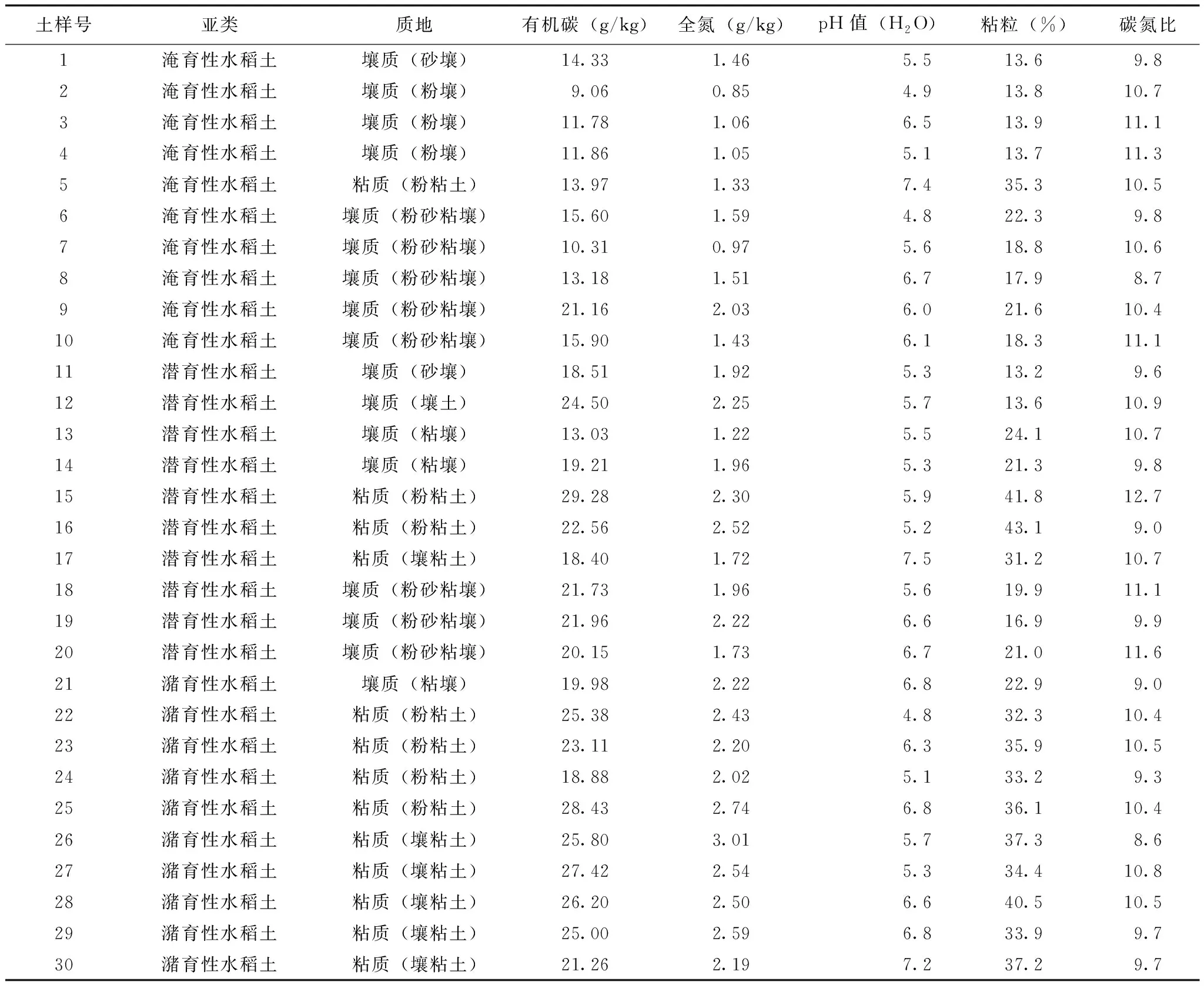
土样号亚类质地有机碳(g/kg)全氮(g/kg)pH值(H2O)粘粒(%)碳氮比1淹育性水稻土壤质(砂壤)14.331.465.513.69.82淹育性水稻土壤质(粉壤)9.060.854.913.810.73淹育性水稻土壤质(粉壤)11.781.066.513.911.14淹育性水稻土壤质(粉壤)11.861.055.113.711.35淹育性水稻土粘质(粉粘土)13.971.337.435.310.56淹育性水稻土壤质(粉砂粘壤)15.601.594.822.39.87淹育性水稻土壤质(粉砂粘壤)10.310.975.618.810.68淹育性水稻土壤质(粉砂粘壤)13.181.516.717.98.79淹育性水稻土壤质(粉砂粘壤)21.162.036.021.610.410淹育性水稻土壤质(粉砂粘壤)15.901.436.118.311.111潜育性水稻土壤质(砂壤)18.511.925.313.29.612潜育性水稻土壤质(壤土)24.502.255.713.610.913潜育性水稻土壤质(粘壤)13.031.225.524.110.714潜育性水稻土壤质(粘壤)19.211.965.321.39.815潜育性水稻土粘质(粉粘土)29.282.305.941.812.716潜育性水稻土粘质(粉粘土)22.562.525.243.19.017潜育性水稻土粘质(壤粘土)18.401.727.531.210.718潜育性水稻土壤质(粉砂粘壤)21.731.965.619.911.119潜育性水稻土壤质(粉砂粘壤)21.962.226.616.99.920潜育性水稻土壤质(粉砂粘壤)20.151.736.721.011.621潴育性水稻土壤质(粘壤)19.982.226.822.99.022潴育性水稻土粘质(粉粘土)25.382.434.832.310.423潴育性水稻土粘质(粉粘土)23.112.206.335.910.524潴育性水稻土粘质(粉粘土)18.882.025.133.29.325潴育性水稻土粘质(粉粘土)28.432.746.836.110.426潴育性水稻土粘质(壤粘土)25.803.015.737.38.627潴育性水稻土粘质(壤粘土)27.422.545.334.410.828潴育性水稻土粘质(壤粘土)26.202.506.640.510.529潴育性水稻土粘质(壤粘土)25.002.596.833.99.730潴育性水稻土粘质(壤粘土)21.262.197.237.29.7
注:土壤质地分类采用国际制标准。
1.2 土壤有机氮矿化培养试验
土壤有机氮矿化培养采用改进的淹水密闭培养-间歇淋洗法[14]。称取过2 mm筛的风干土样10 g置于容积为80 mL的离心管中,每一土壤设置3管,加入25 mL蒸馏水,保持淹水密闭状态在30℃下进行培养。在培养0、3、7、14、21、28、42、56、70、90、112 d时取重复土壤样品按照文献[15]所述方法淋洗并测定土壤矿质氮及计算土壤累积矿化氮。土壤有机氮矿化过程用改进的二阶指数模型进行拟合,公式为:
Nt=Nd(1-e-kdt)+Nr(1-e-kr t)
(1)
式中:Nt——t时间内累积净矿化氮量,mg/kg;
Nd、Nr——土壤易、难矿化氮库矿化势,mg/kg;
kd、Kr——土壤易、难矿化氮库一级反应速率常数,mg/(kg·d);
t——培养时间,d。
式(1)中,不同土壤间kd、Kr均取固定值,分别为0.173、0.012 mg/(kg·d),该值的确定方法参照文献[16]。kd、Kr取固定值的缘由在于:一方面,不同土壤间常规二阶指数模型拟合的kd、Kr值差异均较小;另一方面,取值固定可消除常规二阶指数方程中Nd和kd、Nr和kr均为可变参数且取值结果相互影响的缺点,可获取更真实且易比较的Nd和Nr拟合值[16-17]。土壤氮素矿化势(No)值为Nd和Nr模型拟合值之和。
1.3 土壤有机氮组分测定
土壤有机氮组分用Bremner[18]提出的热无机酸水解法测定,即样品中加入6 mol/L HCl,在120℃下封管水解12 h后冷却、过滤并用少量蒸馏水多次淋洗残渣,酸解液中酸解氮、氨态氮、氨态氮+氨基糖氮、氨基酸氮含量分别用凯氏定氮法、氧化镁蒸馏法、pH 11.2的磷酸盐-硼酸盐缓冲液蒸馏法、茚三酮氧化磷酸盐-硼酸盐缓冲液蒸馏法测定;酸未解氮、氨基糖氮、酸解未知氮含量用差减法求得。计算各有机氮组分占全氮比例(%)。
1.4 数据处理
选择淹水培养7、28、112 d土壤累积矿化氮量(分别用Nmin-7、Nmin-28、Nmin-112表示)及No表征土壤可矿化氮。利用SPSS 16.0软件中的Pearson相关分析、多元线性回归、通径分析程序分析土壤有机氮组分与可矿化氮的关系。Nmin-7、Nmin-28、Nmin-112及土壤有机氮组分含量的表达形式为“平均值±标准误”。
2 结果与分析
2.1 土壤有机氮组分含量及其比例
即使理化性质存在较大差异,各试验土壤均以酸解氮构成有机氮的主体,不同土壤酸解氮含量变幅为545.7~2 276.9 mg/kg(表2),占土壤全氮的比例为58.6%~83.8%,均值达72.4%,而酸未解氮占土壤全氮的比例仅达16.2%~41.4%(图2)。从不同类型水稻土间酸解氮含量变化来看,其值在淹育性、潜育性和潴育性水稻土间的变化范围分别为545.7~1 276.9、962.4~1 759.2和1 523.8~2 276.9 mg/kg,总体上依潴育性水稻土、潜育性水稻土、淹育性水稻土的次序明显降低。
表2显示,酸解氮中,各土样均以氨基酸氮含量最高,以氨基糖氮含量最少,氨态氮和未知氮含量介于其间;计算得知试验土壤间各酸解有机氮组分含量变异系数达34.3%~41.2%,具有中等变异性。不同酸解有机氮组分占全氮含量比例的差异亦较大,氨基酸氮、氨基糖氮、氨态氮和未知氮占全氮的比例分别为25.6%~43.1%、2.6%~9.0%、11.9%~22.3%和8.0%~25.3%(图2)。
2.2 土壤可矿化氮变化及其与理化性质相关性
不论水稻土发育类型是否变化,土壤累积矿化氮量均随培养时间延长而逐渐增加,但有机氮在培养初期矿化相对更为迅速,培养期间各发育类型水稻土包含10个土样平均的累积矿化氮量始终以潴育性水稻土最高,潜育性水稻土次之,淹育性水稻土最低(图3)。进一步分析不同培养期(培养7、28和112 d)土壤累积矿化氮量变化显示,随培养时间延长,供试土壤间Nmin-7、Nmin-28和Nmin-112值变幅依次增大,但其变异系数却逐渐降低(表3)。改进的二阶指数模型对各供试土壤培养112 d的氮素矿化过程均有很好的拟合效果(R2=0.993~0.999,P<0.001)(表3),同一土壤No拟合值较Nmin-112值提高8.6%~30.6%,No值在不同类型水稻土间总体表现出潴育性水稻土高于潜育性水稻土,更高于淹育性水稻土的变化特征。
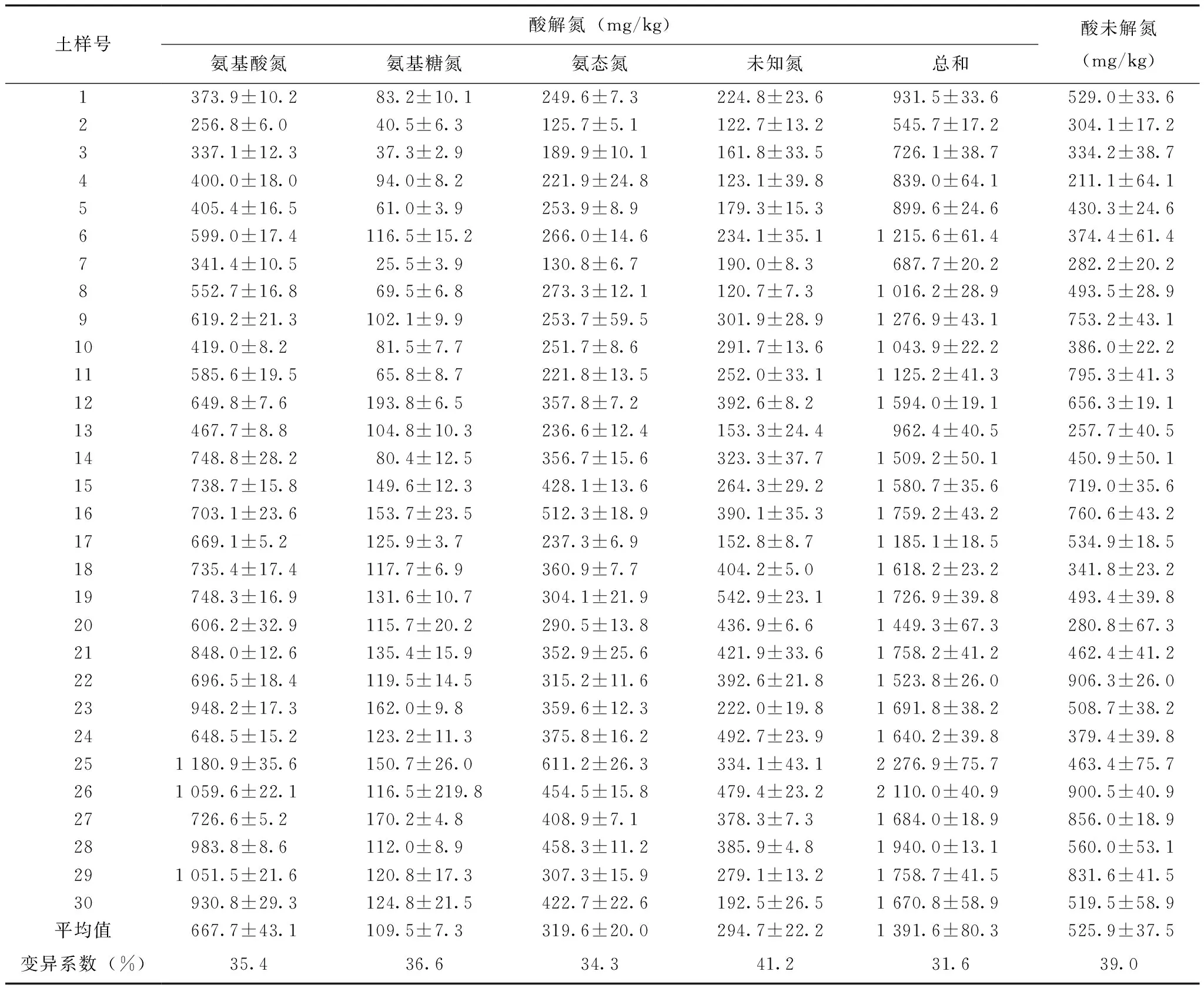
表2 土壤各有机氮组分含量

图2 土壤有机氮组分占全氮比例
分析Nmin-7、Nmin-28、Nmin-112、No值与土壤基本理化性质间的相关关系(表4),结果显示,Nmin-7、Nmin-28、Nmin-112及No均与土壤有机碳、全氮和粘粒含量呈极显著正相关关系(P<0.01),
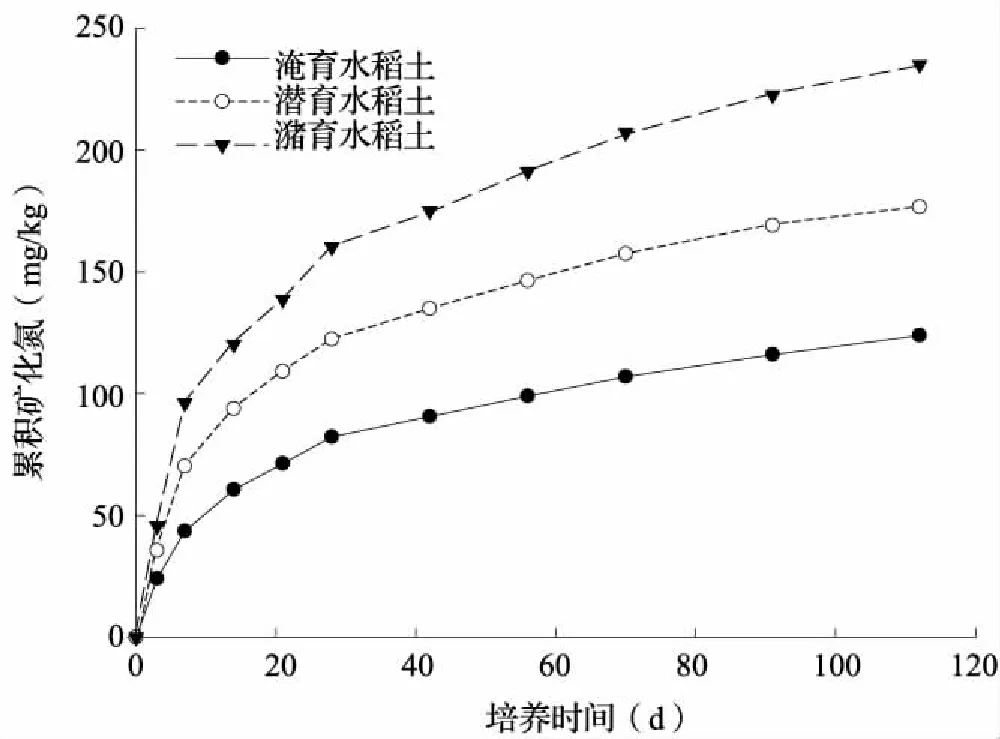
图3 各发育类型水稻土包含10个
但与土壤pH值和碳氮比则不具显著相关关系(P>0.05),显示土壤pH值和碳氮比的变异对供试土壤有机氮矿化无明显影响。

表3 土壤可矿化氮含量
注:R2表示利用改进的二阶指数模型拟合土壤有机氮矿化过程的拟合决定系数,***表示P<0.001显著水平,下同。

表4 土壤可矿化氮与基本理化性质间的Pearson相关系数
注:**表示P<0.01显著水平,下同。
2.3 土壤有机氮组分与可矿化氮的关系
土壤可矿化氮与有机氮组分间的相关分析结果显示(表5),Nmin-7、Nmin-28、Nmin-112及No均与各酸解氮组分呈显著正相关关系(R=0.427~0.858,P<0.05),但与酸未解氮则无显著相关关系(R=0.253~0.318,P>0.05),说明相对于酸未解氮,土壤酸解氮与可矿化氮的关系更为密切。酸解氮组分中,氨基酸氮与可矿化氮间的相关系数始终最高,氨态氮次之,氨基糖氮再次之,酸解未知氮相对最差。
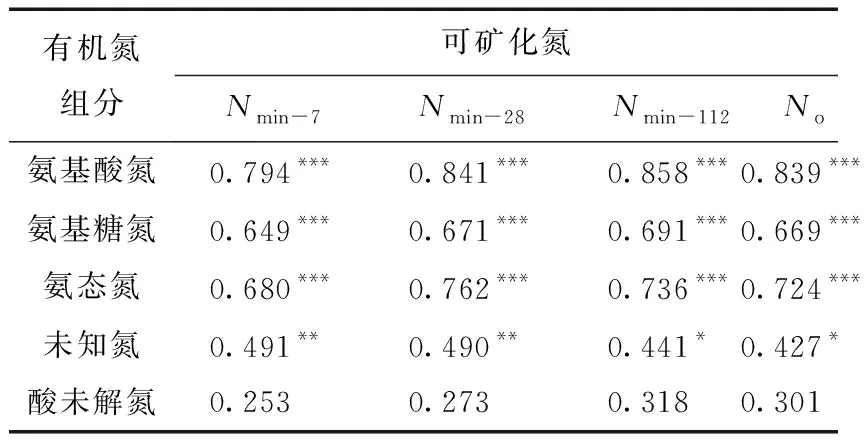
表5 土壤有机氮组分与可矿化氮间的Pearson相关系数
注:*表示P<0.05显著水平。
有机氮各组分间相互关联,使得各酸解氮组分与可矿化氮间的显著相关关系并不能说明前者对后者的真实贡献及贡献大小。为此,进一步进行可矿化氮与酸解氮组分间的多元逐步回归分析,结果显示(表6),酸解氮中,始终只有氨基酸氮进入逐步回归方程,其偏回归系数始终均为正值且逐渐增大,方程决定系数(R2)亦均达0.001显著性水平,表明氨基酸氮对土壤可矿化氮具有稳定且重要的贡献,是后者的主要来源。

表6 土壤酸解氮组分与可矿化氮逐步回归模型输出结果
注:R2表示可矿化氮与酸解氮组分间多元逐步回归分析的方程拟合决定系数。
酸解氮中,氨基糖氮、氨态氮和未知氮与可矿化氮高度相关,但并未能进入逐步回归方程中。因此,需进一步借助通径分析量化土壤各酸解氮组分对可矿化氮的贡献。No表征土壤可矿化氮供应容量,在对其值进行正态性检验后显示,Kolmogorov-Smirnov值为0.455,显著水平(Sig.)=0.986>0.05,表示No值服从正态分布,对其与土壤酸解有机氮组分间的关系进行通径分析。表7显示,虽然各酸解有机氮组分均对No有一定贡献,但其所起直接作用(直接通径系数)和通过其它酸解氮组分所起的间接作用(间接通径系数)却有很大差异。氨基酸氮对No所起的直接作用(0.704)最大,且达P<0.001显著水平,而其通过其它酸解氮组分对No所起的间接作用(-0.028~0.147)均较小,说明氨基酸氮对可矿化氮有显著的直接贡献意义;相反,其它各酸解氮组分对No贡献所起的直接作用均很小,甚至无直接作用(-0.055~0.226),远远低于其通过其它酸解氮组分所起的总间接作用(0.443~0.706)。进一步分析得知,氨基糖氮、氨态氮、未知氮对No的影响均以通过氨基酸氮所起的间接作用(0.353~0.574)最大和最为明显,说明这些组分各自与可矿化氮的良好关系,主要在于其通过其它酸解氮组分尤其是氨基酸氮所起的间接作用。决策系数反映解释变量(酸解氮组分)对响应变量No的综合影响效应[19],氨基酸氮、氨基糖氮、氨态氮的决策系数均为正值且依次减小,说明这些组分均对No起增进作用,且氨基酸氮是主要决策变量;相反,未知氮的决策系数为-0.050,其对No起限制作用。

表7 土壤酸解有机氮组分与氮矿化势(No)间的通径分析结果
注:∑代表各酸解氮组分通过其它酸解氮组分对No的总间接通径系数。
3 讨论
3.1 可矿化氮变化及其土壤控制因子
土壤可矿化氮量是指导农业生产施氮实践的重要参数,其变化反映一定时期内植物可利用态土壤有机氮数量差异[20]。本研究中,培养7、28、112 d条件下,不同土壤间累积矿化氮量变异系数依次降低,其主要原因可能在于:一、不同土壤间氮矿化速率及其差异均随培养时间延长而降低;二、风干土壤氮素矿化存在“干土效应”[21],即风干过程中死亡原生生物体的快速矿化及其对微生物活性的促进作用,导致土壤氮素在培养初期具有更高的矿化速率及矿化变异性。事实上,多数农耕土壤时常经历干湿交替过程,土壤氮素矿化的干土效应在自然状态下亦难以完全避免[16]。不同土壤氮素矿化的干土效应并不一致,但风干过程并不会改变不同土壤间氮素矿化能力的相对顺序[22]。因此,土壤氮素矿化研究常使用风干土壤。No表征土壤可矿化氮供应容量[16]。本研究显示,洞庭湖区不同亚类水稻土No值总体上以潴育性水稻土、潜育性水稻土、淹育性水稻土的次序依次降低,其原因在于潴育性水稻土相对于潜育和淹育性水稻土而言,保持了更好的土壤水分状况,这有利于增强土壤氮循环微生物活性和养分有效性[23]。
土壤可矿化氮变化受许多因素影响,在环境相同情形下,不同土壤间可矿化氮量的差异,主要受其理化特性的控制[24],有机质、碳氮比、质地及pH值等被认为是影响有机氮矿化的重要土壤因子[25]。在一定范围内,土壤碳氮比和pH值往往与可矿化氮分别具负和正相关关系[26-27],这与本研究结果相一致,但未发现土壤可矿化氮量与碳氮比和pH值具显著相关关系,这可能与同区域同类型土壤在相似管理影响下具有较低的pH值和碳氮比值变幅有关。土壤可矿化氮与粘粒含量显著正相关原因在于土壤新形成的团聚体富含易矿化氮素且其易与粘粒结合[27]。土壤可矿化氮与有机碳和全氮含量亦呈极显著正相关(表5),这是因为土壤有机碳和全氮是可矿化氮的源泉,先前的很多研究证实二者同土壤可矿化氮显著正相关[22,24]。然而,鉴于土壤有机碳和全氮含量代表着氮素矿化的理论极限值,其反映土壤供应可矿化氮能力差异只能达到半定量水平[22],因此分析有机碳(氮)组分及其质量与土壤可矿化氮间的关系对深入理解土壤矿化供氮特征及氮素有效性更具实际意义[24]。
3.2 土壤有机氮组分及其与可矿化氮关系
有机态氮化学形态复杂,其各组分含量受一系列自然和人为因素的综合影响。本研究各土壤酸未解氮含量均明显低于酸解氮含量,但亦有研究显示不同轮作模式及施氮量条件下,黄棕壤有机氮均以非酸解氮为构成主体[28],这说明土壤有机氮的酸解效率可能更直接客观地决定于土壤成土特性而非人为活动的干扰。鉴别酸解有机氮组分是深入理解土壤有机氮生物有效性的基础,氨基酸氮是酸解氮中主要可鉴别的含氮化合物,本试验各土壤酸解氮中均以氨基酸氮组分含量最高,而北方一些旱作土壤呈现氨态氮和氨基酸氮含量基本相当[29]或以氨态氮含量最高[30]的特征,这种差异的原因,一方面可能是由于北方气候温暖区土壤粘土矿物多以2∶1型矿物为主,其具备很强的固铵能力[31],而固定态铵是酸解氨态氮的主要来源之一,且在酸解条件下有较高的释放效率[32];另一方面可能是稻田和旱地水热条件的差异影响土壤有机氮组分间相互转化及其分配格局。供试土壤全氮在不同酸解组分中的分配特征与赵丽等[33]的研究结果一致,但计算获得土壤全氮中各有机氮组分比例的变异系数值仅为14.0%~29.7%,较相应组分含量变异系数值降幅达26.7%~60.9%,反映出相似管理条件下同类土壤有机氮素在不同组分间的分配具有一定的共性。
有机态氮是土壤矿质氮的源和库,有机氮分组为揭示土壤不同形态有机氮与可矿化氮间的亲疏关系提供技术支持[4]。简单相关分析揭示土壤有机氮组分与可矿化氮间的平行关系,本研究发现仅酸未解氮未与可矿化氮具显著正相关关系(P>0.05),这与酸未解氮主要以杂环态存在于高度缩合且稳定性较高的腐殖质组分中有直接关系[34]。酸解氮中,未知氮代表相对不易矿化的土壤酸解氮形态[34],多数情况下其有效性较低[35]。本研究中,与其它酸解氮组分进行比较,未知氮表现出与可矿化氮具更弱的正相关关系,且达显著水平(P<0.05)。酸未解氮对No具抑制作用,但有研究显示该组分氮与可矿化氮的关系似乎与土壤淹水培养时长有紧密关联[9]。氨基酸氮与可矿化氮的正相关性最好,且是唯一进入到逐步回归方程的酸解氮组分,这充分说明氨基酸氮对试验土壤可矿化氮供应有重要意义。Lu等[12]研究显示黄土高原旱地土壤淹水条件下以酸解氨态氮对土壤可矿化氮的贡献最大;梁国庆等[36]对石灰性潮土的研究表明,植物吸收态可矿化氮主要来自氨基酸态氮,其次是氨态氮,氨基糖态氮的贡献程度最低;彭银燕等[11]则认为酸解未知氮对不同施肥和地下水位影响下的红壤性水稻土可矿化氮的贡献最大。不同研究结果的差异可能与气候条件、土壤类型及耕作施肥等因素综合影响有关。氨基酸氮与Nmin-7、Nmin-28、Nmin-112建立的逐步回归方程的决定系数(R2)依次增大,表征随土壤氮素矿化作用的增强,氨基酸氮的贡献增大[9]。
土壤有机氮各组分含量的消长及其对土壤可矿化氮的贡献特性,以一种组分间相互转化、相互制约或促进的复杂态势存在,通径分析可在相关分析基础上更精确地解析土壤有机氮组分与可矿化氮间的因果关系[12]。本研究中,虽然酸解氨态氮含量远高于氨基糖氮,二者与可矿化氮间的相关性几无差异,但通径分析中二者间对No的决策系数却存在明显差异,这在一定程度上说明酸解氮组分对可矿化氮的贡献大小与其数量的多寡没有必然联系;酸解氨态氮来源复杂,其中某些来源化合物因具有相对氨基酸氮和氨基糖氮更高的化学稳定性而降低其结合氮素的生物直接可利用性[4]。氨基酸氮对No具有最大的直接作用和最高的决策系数,对试验土壤可矿化氮的综合影响作用最大。之前有研究报道,酸解条件下,碱性氨基酸中的非α-氨基氮和脯氨酸中的氨基氮均不能用茚三酮氧化-磷酸盐-硼酸盐缓冲液蒸馏法测得[34],这些氨基酸态氮被归入到氨态氮或酸解未知氮中,若考虑这些因素,氨基酸氮可能对土壤可矿化氮或植物可利用氮具有更大的贡献[36]。尽管如此,应注意到多种有机氮组分可同时对可矿化氮有贡献,其贡献差异受有机氮组分间相互转化过程的深刻影响。对农业土壤而言,频繁的人为活动对土壤有机氮素形态转化的影响更为强烈和复杂。因此,加强多因素综合影响下土壤有机氮素周转特性研究应是深入理解土壤有机氮组分对可矿化氮的贡献演变及其有效性的重要基础。
4 结论
酸解氮是洞庭湖区双季稻田轮作水稻土有机氮素的主要存在形态,其各组分含量及分配比例以氨基酸氮为最高、氨态氮和酸解未知氮次之,氨基糖氮为最低;不同类型水稻土间酸解氮含量和氮矿化势总体上依潴育性水稻土、潜育性水稻土、淹育性水稻土的次序依次降低。土壤有机碳、全氮及粘粒含量均与土壤可矿化氮间呈显著正相关关系(P<0.01),是控制土壤有机氮矿化的重要理化因子。土壤酸解氮与可矿化氮关系密切,其中,氨基酸氮是对可矿化氮有直接重要贡献的组分,是土壤可矿化氮的主要来源,氨基糖氮、氨态氮和酸解未知氮各自对土壤氮矿化势的影响则主要在于其通过其它酸解氮组分尤其是氨基酸氮所起的间接作用。酸解氮中除未知氮外的其它组分均对土壤氮矿化势起增进作用,提升有机氮中这些组分的分配比例有利于增加土壤可矿化氮供应容量。
