不同天然居群小檗种子萌发障碍因子研究
2018-11-02李林瑜方紫妍艾克拜尔毛拉
李林瑜 方紫妍 艾克拜尔·毛拉 周 龙* 陆 彪
(1.新疆农业大学林学与园艺学院,乌鲁木齐 830052; 2.新疆特克斯县林业局,特克斯县 835500)
种子是大多数木本植物的主要再生手段,它们是植物遗传物质代代相传的的传递系统,成熟的种子在休眠状态下,能够忍受大多数植物不能忍受的不利条件和环境胁迫[1]。许多植物种子的成熟伴随着休眠状态的诱导,种子休眠是抑制植物种子发芽以便等待更有利条件萌发的一种机制[2]。层积是最有效的解除休眠的方法,湿冷或低温层积被广泛应用于打破植物种子休眠和提高其发芽率[3~4]。潘睿等[5]对极度濒危植物大果木莲种子休眠与萌发特性进行研究,结果发现,大果木莲种子具吸水性,胚未完全发育,属于浅度简单型生理休眠,在20/10℃下,1 000 mg·L-1浓度的GA3可以有效打破种子休眠,种子冷层积60 d的萌发率最高。刘延文等[6]研究了不同温度,不同浓度赤霉素处理下蓝靛果忍冬种子的萌发特性,结果发现,赤霉素浓度为100 mg·L-1时,其种子发芽率、发芽指数和发芽势最高;20℃是蓝靛果忍冬种子萌发的最适宜温度。
小檗属(BerberisL.)植物是小檗科(Berberidaceae)最大的属,以灌木和小乔木为主,大多具有冠形优美、枝蔓多姿、花朵清香、果色亮丽等特质,具有较高的观赏价值,是绿化城市的优良材料之一[7~8]。此外,小檗属植物全体都含有小檗碱,入药可燥湿、解毒泻火,在民族医药方面已得到普遍应用和重视[9]。新疆西天山野果林内分布有大量的野生小檗资源,小檗种群在整个流域的地理气候,生物多样性和生态防御体系中起着非常重要而不可替代的作用[10]。库尔班江等[11]曾对伊犁河流域红果小檗资源分布、形态特征及生态条件等进行综合调查。张婷婷[12]和孙颖平[13]等曾对新疆黑果小檗根部化学成分及薄层鉴别方法进行研究。野外调查发现,自然条件下小檗结实量大,但实生苗数量少,野外小檗多以根蘖进行繁殖,导致小檗种群数量及分布范围渐减,而有关西天山野果林野生小檗种子萌发障碍影响因素的研究报道甚少。本研究以西天山野果林3个自然居群内野生小檗种子为试验材料,对不同居群野生小檗种子的生物学特性、萌发特性进行研究,期望为野生小檗的引种、繁殖及育种工作提供技术支持和科学依据。
1 材料与方法
1.1 材料
供试小檗果实均采于西天山野果林不同居群内的小檗自然种群,果实采回后,清洗果皮及果肉,种子自然晾干后,存于4℃冰箱保存备用。发芽实验于2017年10月~2018年1月在新疆农业大学实验室进行。种子分类及种源地自然概况见表1。
1.2 方法
1.2.1 种子形态观测
从自然风干的4组小檗果实中,每组随机选取20个完整果实,剥开并记录每个完整果实中的种子数量。每组随机选取20粒饱满的小檗种子测量种子大小,种子大小以游标卡尺测量的纵横径表示,并用目测法观察种子颜色、光滑度,质感等。采用百粒法[14]测种子千粒重,从每组小檗种子中随机数取5份(100粒/份)种子,分别称重记录,取5份种子平均重量乘以10即为种子千粒重。
1.2.2 种子活力测定
采用TTC法[15]测定种子活力,每组随机选取30粒小檗种子,在25℃水中浸种24 h,待种子吸胀后用刀片沿种子胚中心线纵切为两半,将其中一半置于培养皿中,加入适量的0.5% TTC溶液,以淹没种子为度。然后置于35℃恒温箱中染色1 h,凡胚全部被染为红色的为具有生活力的种子,重复3次,观察并记录结果。

表1 霍城、新源和特克斯小檗种源地自然概况
1.2.3 种子吸水率测定
从4组小檗种子中,每组称取1 g小檗种子,加蒸馏水50 mL,放置于室温条件下,每隔2 h取出,用滤纸擦干种子表面水分称重,12 h后每12 h取出称重,直到重量不变为止,最后计算种子吸水率,3次重复。种子吸水率是种子增加的重量与风干种子重量的百分比。
1.2.4 种子萌发特性的测定
分别将4组小檗干种子洗净用清水浸泡24 h与湿度50%的干净河沙混匀(体积比:沙量∶种量=4∶1)于4℃条件下层积。隔15 d从每组层积种子中取出80粒不剥皮种子和80粒种子剥除种皮(用小刀将种子的头部划开,剥去外种皮,保留完整的子叶),分别分为四组进行不同处理:CK清水浸泡2 h;浓度为100 mg·L-1GA3溶液浸泡2 h;浓度为200 mg·L-1GA3溶液浸泡2 h;④浓度为300 mg·L-1GA3溶液浸泡2 h。处理后种子用蒸馏水清洗两遍,进行发芽试验,统计其发芽率,重复3次。
层积前种子用3%硫酸铜溶液消毒1 h,沙子经高压灭菌锅120℃灭菌30 min。发芽实验采用培养皿发芽法,将小檗种子播于铺有5层滤纸6层纱布的培养皿中,每皿20粒,在25℃恒温条件下进行发芽试验。以胚根露出种皮1 mm为发芽标准,以发芽率达到70%作为种子解除休眠的阈值。
2 结果与分析
2.1 种子形态观测及活力测定
不同天然居群小檗种子的形态,大小各有差异(表2)。霍城、新源和特克斯居群黑果小檗每粒完整果实含种子数为3~4粒,红果小檗果实含种子数为2~3粒;黑果小檗种子呈棕红色,红果小檗种子为棕黄色。霍城居群黑果小檗种子最大,其千粒重均值为18.49 g、纵径均值为5.64 mm、横径均值为2.76 mm均大于新源、特克斯居群黑果小檗种子。红果小檗种子较小,千粒重均值为7.72 g、纵径均值为3.87 mm、横径均值为1.81 mm均小于黑果小檗种子。不同居群小檗种子活力无显著性差异,特克斯居群黑果小檗种子活力最高为79.67%。
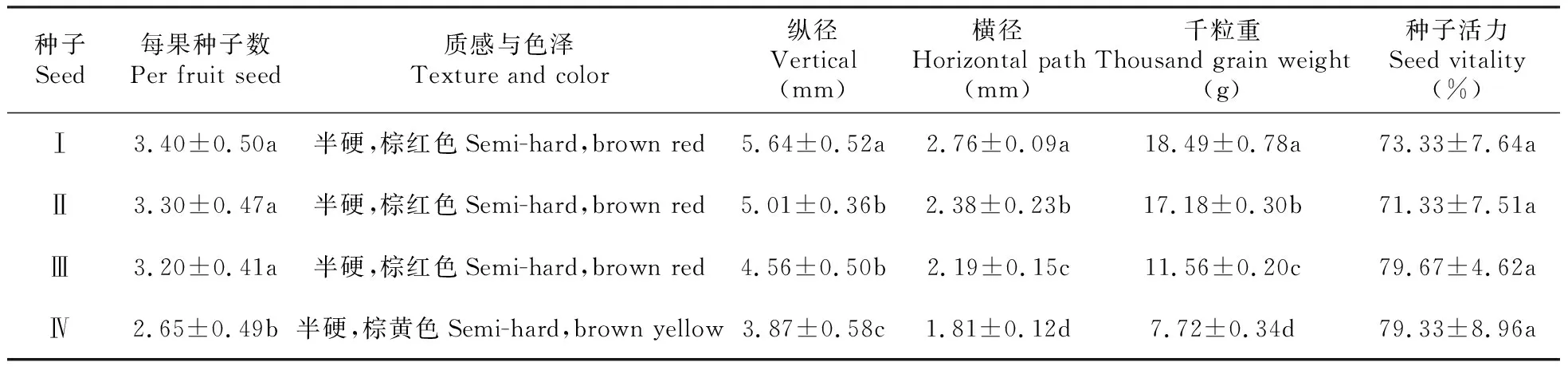
表2 不同天然居群小檗种子形态观测
注:表格中数字均为平均值±标准误,同列不同字母之间表示差异显著(P≦0.05),下同。
Note:The figures in the table are the mean±standard error,and the difference between the letters in the same column indicates that the difference is significant(P≦0.05),the same as below.

图1 黑果小檗与红果小檗 A.黑果小檗果实;B.黑果小檗种子;C.红果小檗果实;D.红果小檗种子Fig.1 Black fruit Berberis and red fruit Berberis A. Fruit of the black-fruit Berberis; B. Seed of the black-fruit Berberis; C. Fruit of red-fruit Berberis; D. Seed of red-fruit Berberis
2.2 种子吸水特性
根据图2可知,4种野生小檗种子的吸水率均表现出逐渐增加的趋势。种子吸水前12 h内,4种小檗种子吸水率增幅最大值均出现在吸水2 h时,霍城居群黑果小檗种子吸水2 h时吸水率为13.93%,其次为红果小檗种子13.90%,新源居群和特克斯居群黑果小檗种子较低分别为7.92%和6.08%。黑果小檗种子与红果小檗种子吸水率差异不明显,其种皮透水性相似。
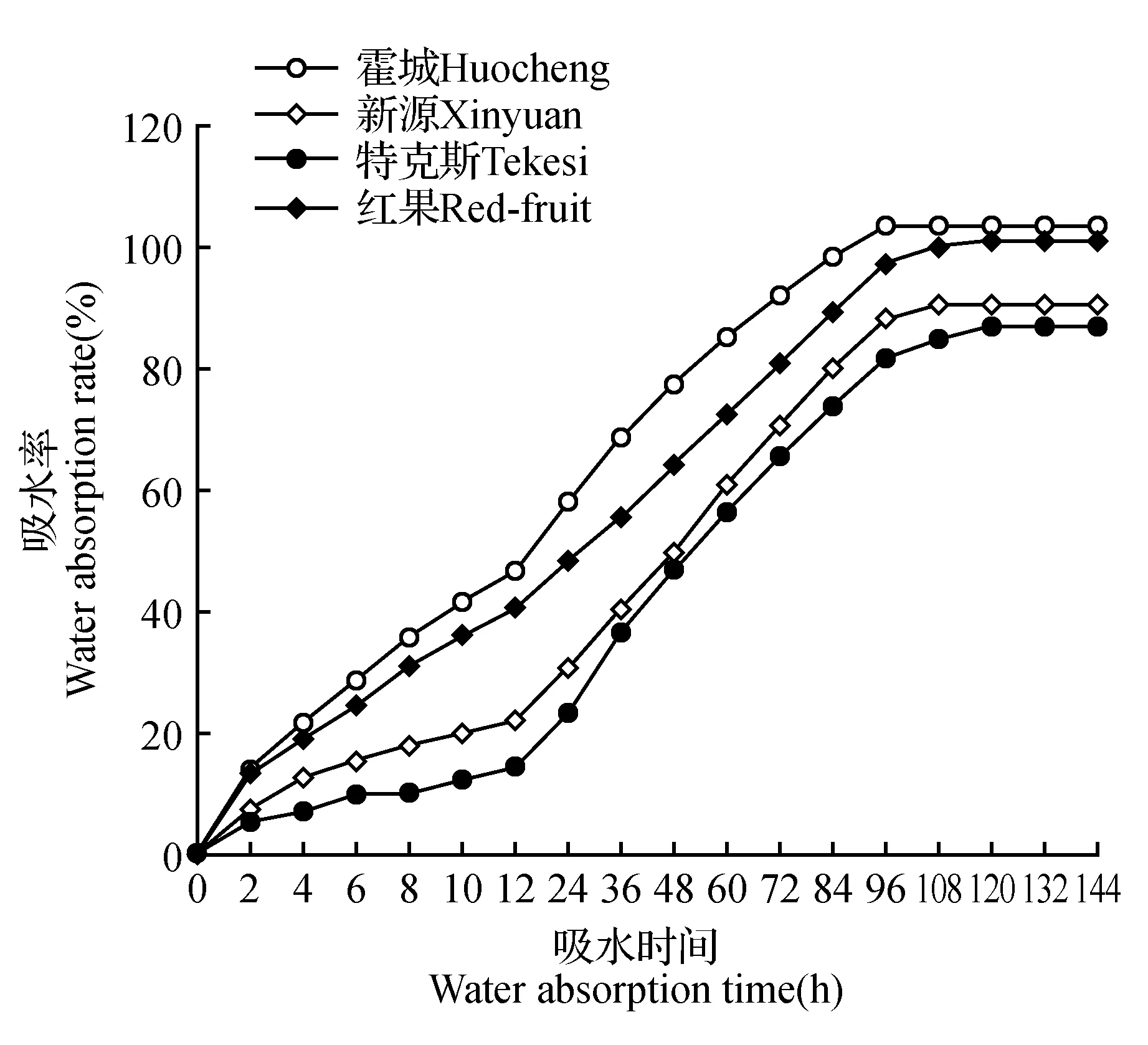
图2 不同天然居群小檗种子吸水特性Fig.2 Water absorption characteristics of seeds of different natural populations
2.3 种子萌发特性
2.3.1 种皮和层积时间对小檗种子萌发的影响
如表3所示,黑果小檗种子与红果小檗种子萌发特性不同,黑果小檗层积0 d时带种皮和去种皮种子均无萌发,而红果小檗带种皮和去种皮种子不需低温层积均可萌发。Ⅰ、Ⅱ、Ⅲ 3组黑果小檗层积30d以内带种皮种子均无萌发,层积45 d时Ⅰ、Ⅱ、Ⅲ带种皮种子开始萌发,发芽率分别为5.42%、5.83%和8.33%。随层积时间的增加,3组带种皮种子发芽率均平稳上升,层积90 d时Ⅰ、Ⅱ、Ⅲ带种皮种子发芽率分别提高到70.00%、70.00%和72.08%。红果小檗种子层积0 d时带种皮种子发芽率为52.92%,层积30 d时红果小檗带种皮种子发芽率提高到70.42%。
Ⅰ、Ⅱ、Ⅲ 3组黑果小檗层积15 d时去种皮种子,发芽率分别为11.52%、15.42%和17.92%,层积60 d时Ⅰ、Ⅱ、Ⅲ 3组黑果小檗去种皮种子,发芽率分别提高到到70.83%、70.00%和70.42%,种子休眠基本被解除。黑果小檗及红果小檗种子萌发情况及幼苗见图3。
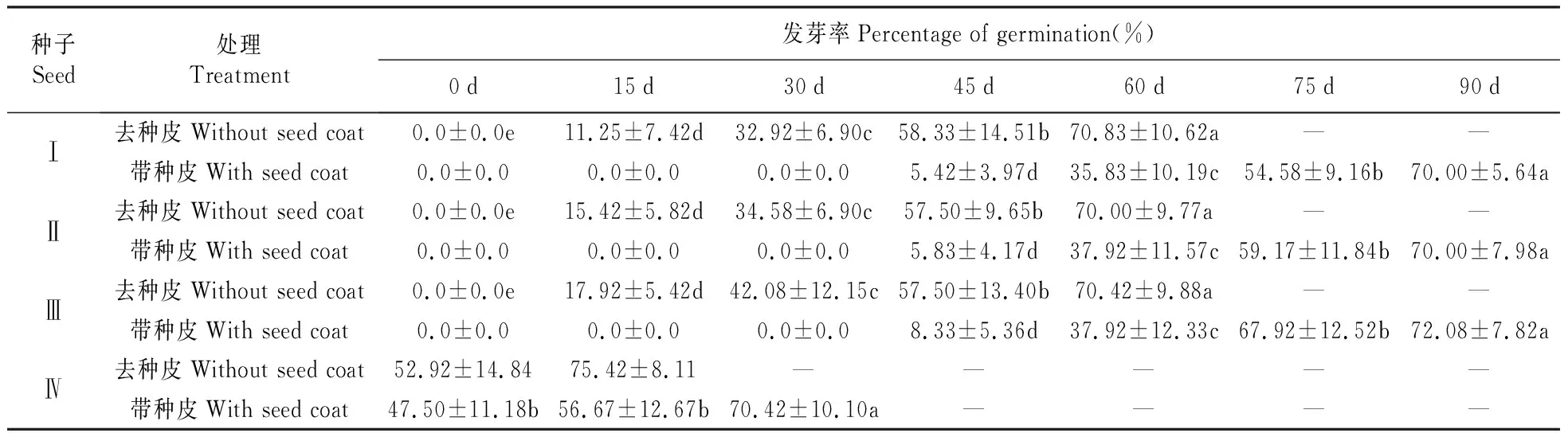
表3 种皮和低温层积对小檗种子发芽率的影响

图3 黑果小檗与红果小檗种子萌发及幼苗 A1.黑果小檗带种皮种子萌发;A2.黑果小檗去种皮种子萌发;A3.黑果小檗幼苗两叶期;A4.黑果小檗幼苗显露第一片特征叶;A5.黑果小檗幼苗;B1.红果小檗带种皮种子萌发;B2.红果小檗去种皮种子萌发;B3.红果小檗幼苗两叶期;B4.红果小檗幼苗显露第一片特征叶;B5.红果小檗幼苗Fig.3 Seed germination and seedling of black and red fruit Berberis A1. Germination of the black-fruit Berberis seed with seed coat; A2. Seed germination of the black-fruit Berberis seed withput seed coat; A3. Two-leaf stage of the black-fruit Berberis seedlings; A4. First characteristic leaf of the black-fruit Berberis seedlings; A5. Seedling of the black-fruit Berberis seedlings; B1. Germination of the red-fruit Berberis seed with seed coat; B2. Seed germination of the red-fruit Berberis seed withput seed coat; B3. Two-leaf stage of the red-fruit Berberis seedlings; A4. First characteristic leaf of the red-fruit Berberis seedlings; A5. Seedling of the red-fruit Berberis seedlings
2.3.2不同浓度GA3处理对小檗种子萌发的影响
由图4可知,不同浓度GA3处理,Ⅰ、Ⅱ、Ⅲ、Ⅳ四组小檗去种皮种子发芽率差异显著,其中GA3浓度为200 mg·L-1时,各组小檗去种皮种子发芽率最高。随着层积时间的增加,各组小檗去种皮种子发芽率逐渐提高,Ⅰ、Ⅱ、Ⅲ 3组黑果小檗层积60 d时去种皮种子用浓度为200 mg·L-1的GA3溶液处理后,发芽率分别为85.00%、83.33%和86.67%,均明显高于GA3浓度为0、100和300 mg·L-1时。红果小檗层积15 d时去种皮种子用浓度为200 mg·L-1的GA3溶液处理后,发芽率可达86.67%,明显高于GA3浓度为0、100和300 mg·L-1时的发芽率66.67%、71.67%和76.67%。
根据图5可知,带种皮黑果小檗种子需要一定时间的低温层积才能萌发,Ⅰ、Ⅱ、Ⅲ组黑果小檗层积45 d带种皮种子均开始发芽,不同浓度GA3处理各组种子后对发芽率的影响明显。层积90 d时,Ⅰ、Ⅱ、Ⅲ 3组黑果小檗带种皮种子用浓度为200 mg·L-1的GA3溶液处理后,发芽率分别达到81.67%、81.67%和86.67%,明显优于其它3组处理。红果小檗层积30 d时带种皮种子用浓度为200 mg·L-1的GA3溶液处理后,发芽率可达85.00%,明显高于GA3浓度为0、100和300 mg·L-1时的发芽率61.67%、71.67%和63.33%。
3 讨论
3.1 种皮对小檗种子萌发的影响
种子的萌发与自身物理特性相关,种皮的不透气性、不透水性、机械阻碍及种皮内含有抑制物质都会引起种子的休眠。本研究发现,Ⅰ、Ⅱ、Ⅲ 3组黑果小檗种子层积15 d时,去种皮种子开始萌发,对层积15 d以内的黑果小檗种子而言种皮并非影响其萌发的主要因素,可能是其种胚中存在萌发抑制物;层积45 d时,带种皮种子开始萌发,种皮成为抑制黑果小檗种子萌发的主要因素,随层积时间的增加,种皮抑制萌发的作用逐渐减弱,层积90 d时黑果小檗带种皮种子发芽率基本达到70%,种皮抑制萌发的作用基本被解除。卢明艳等[16]在研究5种野生无髯鸢尾种子休眠与萌发特性时发现,去种皮能促进溪荪种子的萌发,种皮的机械阻力和种皮中存在萌发抑制物是影响溪荪种子萌发的主要因素。郎思睿等[17]在对香叶树种子休眠与萌发特性的研究中同样发现,香叶树的萌发抑制因素主要来源于种皮的内源抑制物。黑果小檗种子去种皮后萌发速度高于带种皮种子,表明其种皮对黑果小檗种子萌发的抑制作用明显。红果小檗去种皮和带种皮种子不需低温层积即可萌发,发芽率分别为52.92%和47.50%,由此认为种皮对红果小檗种子萌发无明显影响。根据黑果小檗种子与红果小檗种子吸水特性测定结果,可以发现黑果小檗与红果小檗种子均能较快吸胀饱和;对小檗种子进行剥除种皮处理时发现,红果较黑果小檗种子更易于去种皮,黑果小檗种子种皮机械强度比红果小檗稍大。因此推测认为,黑果小檗种子萌发过程中,其种皮的机械抑制作用和存在萌发抑制物可能是其萌发障碍的关键影响因素。
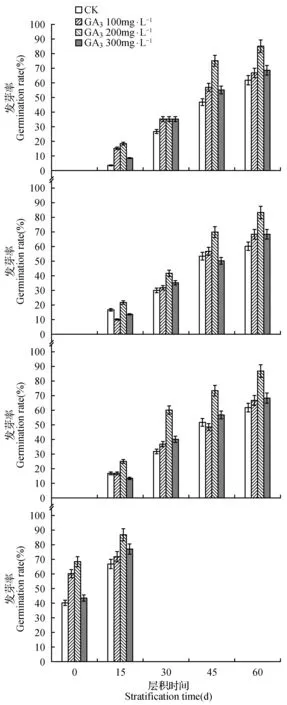
图4 不同浓度GA3处理对不同居群小檗去种皮种子发芽率的影响 A.霍城黑果小檗去种皮种子;B.新源黑果小檗去种皮种子;C.特克斯黑果小檗去种皮种子;D.红果小檗去种皮种子Fig.4 Effect of GA3 treatment with different concentrations on the germination rate of different populations Berberis without seed coats A. Huocheng black fruit Berberis seed without seed coat; B. Xinyuan black fruit Berberis seed without seed coat; C. Tekesi black fruit Berberis seed without seed coat; D. Tekesi red fruit Berberis seed without seed coat
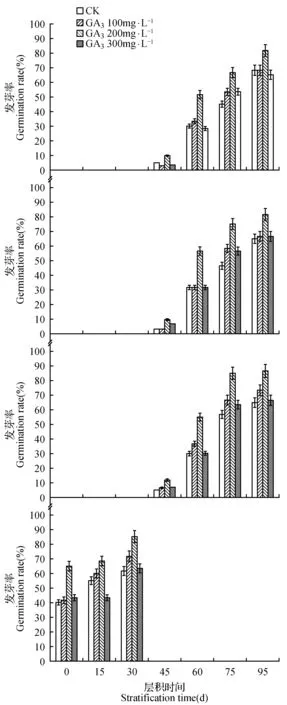
图5 不同浓度GA3处理对不同居群小檗带种皮种子发芽率的影响 A.霍城黑果小檗带种皮种子;B.新源黑果小檗带种皮种子;C.特克斯黑果小檗带种皮种子;D.红果小檗带种皮种子Fig.5 Effect of GA3 treatment with different concentrations on the germination rate of different populations Berberis with seed coats A. Huocheng black fruit Berberis seed with seed coat; B. Xinyuan black fruit Berberis seed with seed coat; C. Tekesi black fruit Berberis seed with seed coat; D. Tekesi red fruit Berberis seed with seed coat
3.2 低温层积对小檗种子萌发的影响
大多数研究者认为低温层积处理可有效打破植物种子休眠,从而促进种子萌发。本试验发现,4℃低温层积处理能够有效解除小檗种子的休眠,随层积时间的增加小檗种子发芽率均逐渐提高。司倩倩等[18]在研究玉铃花种子休眠原因及其内源激素含量变化时同样发现,低温层积可有效打破玉铃花种子休眠,随着层积时间的增加,发芽的种子数逐渐增加。周芝琴等[19]以嵩草、黑穗苔草、藨草和苔草4种莎草科植物种子为材料,研究其种子休眠与萌发特性发现,4种参试植物种子均存在不同程度的生理休眠,低温层积处理对种子萌发的影响因种与层积时间而异。红果小檗种子层积30 d和黑果小檗种子层积90 d时发芽率均在70%以上,种子休眠基本被解除,推测认为随着种子层积时间的改变小檗种皮和胚中萌发抑制物对萌发的影响程度也在改变。低温层积一方面能通过湿润状态逐渐软化种皮,另一方面在层积过程中种子生理指标及内源激素含量也在不断变化,层积催芽能使种子合成生长激素,降解抑制激素。
3.3 不同浓度GA3处理对小檗种子萌发的影响
GA3参与调控种子发芽、茎生长、开花发育、果实成熟等各阶段发育过程,常被用于打破种子休眠,不同植物种子最适宜的GA3浓度不同,低于最适浓度时,GA3浓度越高,越能促进植物生长,高于最适浓度时,抑制作用较明显。本试验结果表明,用浓度为200 mg·L-1GA3溶液对小檗去种皮和带种皮种子进行处理,有利于打破小檗种子的休眠,提高种子的发芽率。唐澄莹等[20]在对珍稀植物北碚榕的种子萌发及繁殖进行研究时发现,用浓度为50~500 mg·L-1GA3溶液浸种4 h后,北碚榕种子发芽率比对照温水浸种显著提高,且发芽率随GA3质量浓度提高呈先升后降变化趋势。翟淑华等[21]用浓度为25、50、75和100 mg·L-1的GA3溶液处理海菜花种子,种子发芽率随GA3浓度升高呈先提高后下降的变化趋势,浓度为75 mg·L-1的GA3溶液处理海菜花种子效果较为理想。本研究用浓度为200 mg·L-1GA3溶液处理小檗种子,其发芽率最高,浓度过高或者过低均会对小檗种子萌发起到抑制作用。适宜浓度的GA3能取代一些种子对低温后熟、光暗和干藏后熟条件的需求,通常可解除种子的浅度生理休眠,外源添加GA3可能增加了小檗种子总GA3的含量,导致种子内GA3/ABA的比率提高,促进小檗种子从休眠或静止状态向萌发过渡。
4 结论
通过对西天山野果林霍城居群、新源居群、特克斯居群黑果小檗以及特克斯居群红果小檗种子萌发特性的研究,发现小檗种子均具有不同程度的生理休眠。对于具浅度生理休眠的红果小檗种子,层积30 d和添加200 mg·L-1GA3处理可有效解除其休眠;对于具有中度休眠的霍城居群、新源居群、特克斯居群黑果小檗种子,剥除种皮、层积90 d和添加200 mg·L-1GA3处理均可有效破除其休眠。
