野生东方田鼠4种成纤维细胞培养上清对日本血吸虫童虫的体外杀伤效果
2018-10-30李海霞汤志宏
吴 侠, 成 钢, 黎 欢, 李海霞, 汤志宏
(1. 南部战区空军参谋部门诊部妇儿科, 广州 510071;2. 湖南文理学院生命与环境科学学院动物健康养殖研究所, 常德 415000)
随着东方田鼠(Microtus fortis)实验动物化、标准化及规范化进程的稳步推进,作为目前重要的天然抗日本血吸虫病实验模型动物,东方田鼠越来越受到广大科研工作者的关注[1,2]。大量研究[3,4]证明,东方田鼠抗日本血吸虫作用是其体内诸多组织与细胞参与渐进性过程,是通过细胞机械性防御和分泌细胞因子及其它活性成分,以多种途径参与其抗虫免疫和防御 。正常小鼠寄生虫感染后可引起大量游离单核细胞、 巨噬细胞和淋巴细胞聚集与活化,产生一系列细胞因子和活性物质进行免疫调节、参与炎症反应及创伤愈合等过程[5]。前期试验已证明东方田鼠不同组织成纤维细胞传代次数是相同的[6-9],为了充分利用不同组织来源的成纤维细胞培养液中活性成分,本试验分别收集处于对数生长期的第三代东方田鼠胚胎和皮肤成纤维细胞,第二代腹腔成纤维细胞,以及第一代肺脏成纤维细胞培养上清液与日本血吸虫童虫共培养,对虫体进行形态学特征比较观察,为进一步从细胞、分子水平深入了解东方田鼠抗日本血吸虫机制,加快和拓宽探寻东方田鼠抗日本血吸虫活性分子提供参考。
1 材料与方法
1.1 动物来源
野生东方田鼠5对捕自衡阳市衡东县新塘镇杨塘河坝,经鉴定均为长江亚种(Mf. calamorum),随机分组,1对/笼,常规方法饲养于湖南文理学院生命与环境科学学院动物科学专业实验室,饲养条件与饲料同昆明小鼠[SCXK(湘)2016-0002]。雌雄合笼后,计算妊娠天数,根据试验需要取不同日龄东方田鼠作为试验用鼠。普通级雄性新西兰白兔(1.5~2.0 kg)由中南大学湘雅医学院实验动物学部提供[SCXK(湘)2017-0002]。 日本血吸虫阳性钉螺购自湖南省血吸虫病防治所。
1.2 主要试剂
DMEM(高糖)培养基(美国 Invitrogen 公司),新生牛血清(NBS杭州四季青生物工程有限公司),胰蛋白酶(上海生工生物工程股份有限公司); 无钙镁PBS溶液由实验室配制消毒,其余常用试剂均为国产分析纯。东方田鼠血清制备采用摘眼球取血法,兔血清制备采用心脏采血法,分别将采集的两种血液放入离心管内,37℃条件下放置15 min,待自然析出血清后,1 000 r/min 离心10 min,吸取上清,-70℃保存备用。
1.3 方法
1.3.1 野生东方田鼠胚胎、皮肤、肺脏及腹腔成纤维细胞原代与传代培养 根据文献[6-8]报道,取妊娠12 d的东方田鼠胎鼠和出生后1 ~3 d 东方田鼠乳鼠皮肤和肺脏组织分别运用胰蛋白酶消化法和组织块贴壁法进行成纤维细胞体外分离、培养。用无菌PBS灌洗成年野生东方田鼠腹腔,离心洗涤细胞沉淀进行腹腔成纤维细胞分离与培养[9]。以含150 mL/L新生牛血清的DMEM高糖培养基培养上述4种细胞, 当原代细胞铺满培养瓶的70%~80%时, 用质量分数0.25%胰蛋白酶消化传代。原代和传代细胞均置于37 ℃, 体积分数5% CO2条件下培养, 定期观察、记录4种细胞的生长状态, 根据试验需要定期收集处于对数生长期的细胞培养上清。
1.3.2 细胞培养上清液的收集与处理 当成纤维细胞处于快速生长的对数期时,即胚胎、皮肤成纤维细胞第三代,腹腔成纤维细胞第二代,肺脏成纤维细胞第1代,小心收集上述细胞培养上清后,2 000 r/min 离心 10 min 取上清, -70 ℃ 保存备用。
1.3.3 日本血吸虫童虫培养 用消毒生理盐水将阳性钉螺 20只冲洗2次,放入三角瓶,在 25 ℃有光照条件下,常规方法逸出尾蚴,用接种环定时多次收集上层尾蚴后,用无菌PBS 1 000 r/min 离心洗涤尾蚴2次,将尾蚴放入48孔细胞培养板,用含体积分数 25%新鲜兔血清的DMEM培养基培养血吸虫尾蚴,于 37 ℃、体积分数5% CO2条件下培养24 h,调整尾蚴密度,待其体外自然转化为血吸虫童虫[8]。
1.3.4 东方田鼠成纤维细胞培养上清杀伤血吸虫童虫试验 当日本血吸虫尾蚴体外自然转化为童虫后,吸除每孔童虫中全部培养基,将收集的4种细胞培养上清液于37 ℃解冻,将上清液转移到含尾蚴的48孔培养板中, 调整尾蚴100±10 尾/孔, 每个样本设立平行 3孔, 以不加其它物质的DMEM 培养基为空白对照, 以含20 % 浓度的东方田鼠血清为阳性对照, 于37 ℃、体积分数5% CO2条件下培养, 96 h内定时观察东方田鼠4种成纤维细胞培养上清体外杀伤日本血吸虫童虫的效果, 统计童虫死亡率[8]。
1.3.5 童虫死活鉴定标准 活童虫一般培养24 h后分叉的尾巴脱落,虫体透明,自由蠕动伸缩,内部器官清晰可辨,美兰染色易着色; 死亡童虫虫体模糊,表膜破损断裂、内容物外泄、镜下观察无立体感,美兰染色一般不着色; 不易辨别死活的童虫以连续镜下观察 30 s不活动判定为死童虫[2,3]。
1.3.6 统计学分析
童虫死亡率数据用表示,统计分析由SPSS11.5软件中One-way ANOVA分析完成,P<0.05为差异具有统计学意义。
2 结果
2.1 成纤维细胞培养上清与血吸虫童虫共培养
试验96 h 内连续观察了野生东方田鼠4种成纤维细胞培养上清与血吸虫童虫共培养结果。培养结果表明, 与空白对照组和胚胎、皮肤成纤维细胞培养上清童虫相比, 东方田鼠腹腔与肺脏成纤维细胞培养上清中童虫死亡率明显增高, 童虫大多在72 h之内呈现死亡特征, 表现为表膜皱缩,缢痕明显; 培养96 h后, 虫体内容物模糊并溢出,虫体周围体壁上常沾附较多泡状物,与添加20%东方田鼠血清的阳性对照组死亡童虫状态略有不同(图1A~F)。
3.2 成纤维细胞培养上清中的童虫死亡率
取野生东方田鼠4种成纤维细胞培养上液与日本血吸虫童虫共培养96 h后,东方田鼠皮肤、胚胎、腹腔和肺脏组织的成纤维细胞培养上清中童虫死亡率,与空白对照和相比,腹腔和肺脏成纤维细胞对日本血吸虫童虫有一定的杀伤效果(P=0.017;P=0.039),详见表1。
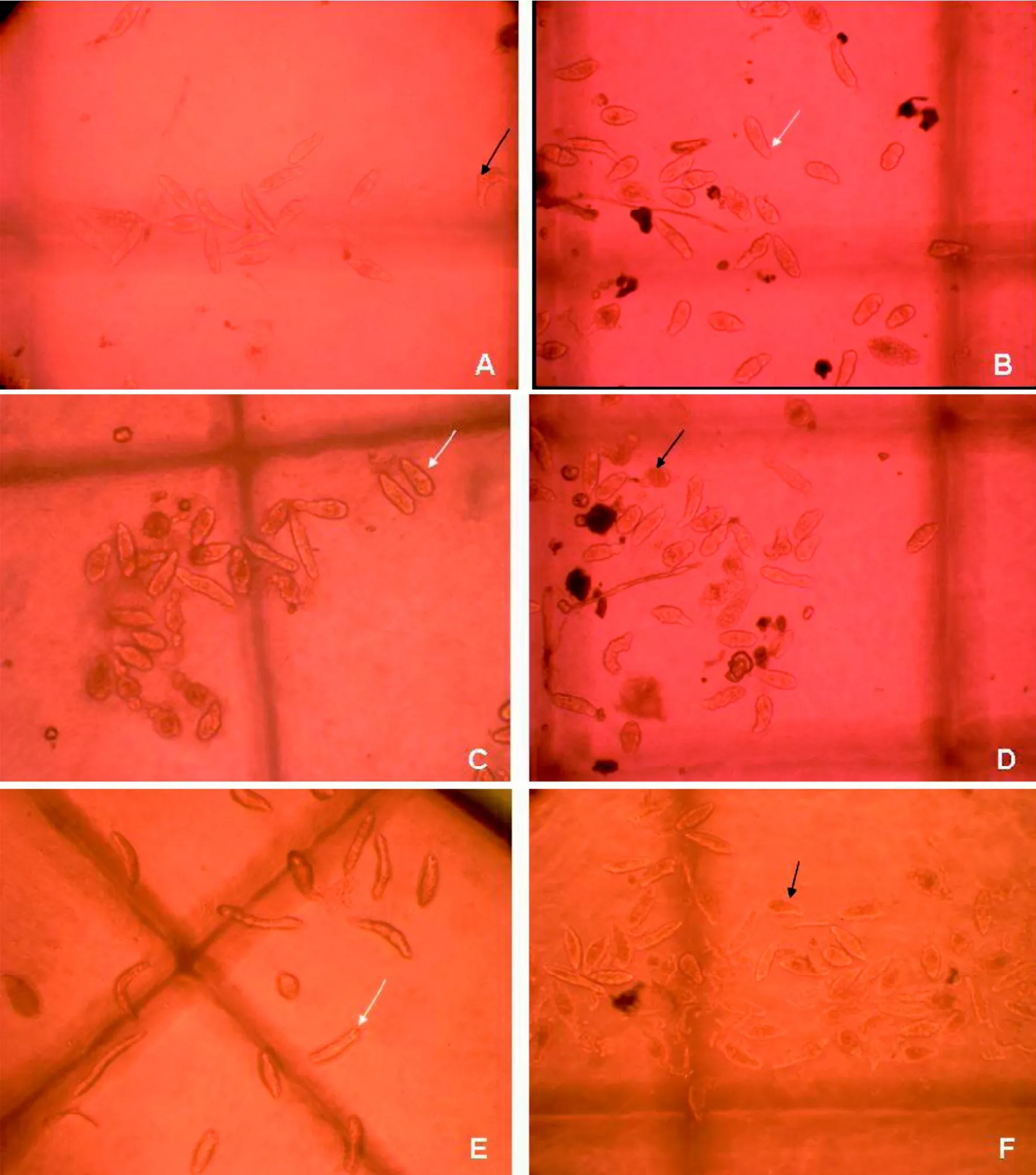
图1 野生东方田鼠4种成纤维细胞培养上清96 h后体外杀伤日本血吸虫童虫效果 (×100)Figure 1 Killing effect of 4 kinds of fibroblasts culture supernatant on schistosomula (×100)
3 讨论
东方田鼠是目前已知鼠类中抗日本血吸虫能力最强的啮齿类哺乳动物[10,11]。贺宏斌等[12]早期研究表明,野生较室内繁殖东方田鼠在日本血吸虫感染后肝脏表面结节大、数量多,消退快,说明野生东方田鼠对日本血吸虫感染应激和免疫反应更大。本课题组前期研究中曾经以室内繁殖东方田鼠细胞培养上清液与血吸虫童虫共培养96 h,并未观察到明显的童虫杀伤作用,可能与添加培养上清液浓度较小有关。为了进一步明确野生东方田鼠细胞培养上清中是否存在抗日本血吸虫分子,以及加快和拓宽探寻东方田鼠抗日本血吸虫活性分子的进程与渠道,试验收集了处于对数生长期的4种成纤维细胞培养上清液,以100%添加量,96 h内连续观察了对血吸虫童虫体外杀伤效果。培养结果表明,与对照相比,腹腔和肺脏成纤维细胞对日本血吸虫童虫有一定的杀伤作用,而且童虫死亡状态与阳性对照组童虫明显不同。成纤维细胞可以向培养液中分泌细胞生长因子、白血病抑制因子等种类繁多的蛋白成分,具有免疫调节、促进损伤组织修复等功能,童虫死亡状态和杀伤效果的不同,是否表明在野生东方田鼠腹腔和肺脏成纤维细胞培养上清中存在能引起血吸虫童虫死亡的抗虫活性分子。为了阐明上述推论,课题组已展开对野生东方田鼠腹腔和肺脏成纤维细胞培养上清中抗性分子的分离和筛选工作,希望对东方田鼠抗日本血吸虫感染机制相关研究提供新的线索。

表 1 野生东方田鼠4种成纤维细胞培养上液与日本血吸虫童虫共培养96 h后结果Table 1 Killing effect of 4 kinds of fibroblasts culture supernatant on schistosomula
