胡麻RIL群体苗期抗旱性状的主基因+多基因遗传分析
2018-10-25王利民张建平谢亚萍牛早霞
王 斌,王利民,张建平,,谢亚萍,牛早霞,赵 利
(1.甘肃省农业科学院作物研究所,甘肃 兰州 730070;2.甘肃农业大学农学院,甘肃 兰州 730070)
胡麻(LinumusitatissimumL.)是北方地区重要的油料作物,胡麻油含有对人体有益的α-亚麻酸、胡麻籽富含木酚素和膳食纤维含量等多种营养保健成分。随着社会经济的发展和生活水平的提高,人们越来越注重食物中营养元素和功能性膳食纤维,胡麻籽独特的营养保健作用,受到人们越来越多的重视。胡麻主要分布在我国西北和华北地区干旱贫瘠的地区,该地区年降水量一般400~500 mm,北部只有200~300 mm,属于干旱半干旱农业生产区,干旱是造成胡麻产量低而不稳的最主要因素[1]。胡麻生产面临着种植效益低,农民种植积极性下降,全国胡麻种植面积持续下滑等诸多问题。改良胡麻品种,提高胡麻抗旱性和产量是胡麻产业持续稳定发展的急需解决的问题。
目前,胡麻抗旱性研究主要针对成株期和萌发期,苗期抗旱性研究比较少。祁旭升等对胡麻种质资源成株期抗旱性进行了评价并进行抗旱指标的筛选[2,3]。罗俊杰等对胡麻主要育成品种成株期抗旱性进行了评价[4,5]。石仓吉通过水、旱地对15个品种的11个性状进行了抗旱性鉴定[6]。何丽等研究了干旱对胡麻现蕾期光合特性及产量的影响[7]。吴文荣,李娜,张延军,姚玉波,王宗胜等对胡麻萌发期抗旱性进行了评价[8-12]。赵利等对胡麻苗期抗旱性的生理指标进行了研究[13],而对胡麻苗期抗旱性相关农艺性状的研究比较少。
作物抗旱性属于复杂的数量性状,受环境影响比较大。有关胡麻抗旱机理和抗旱遗传研究国内外报道比较少,研究基础比较薄弱。盖钧镒、章元明等提出的主基因+多基因混合遗传模型在统计分析植物数量性状的应用比较广泛[14,15]。Hu等用此方法对东乡野生稻BILs群体苗期抗旱性的农艺性状进行了遗传分析[16]。在胡麻上,利用该方法对胡麻农艺、产量和品质性状,胡麻温敏雄性不育产量相关性状,脂肪酸含量和粗脂肪等数量性状进行了研究[17-21]。在胡麻上还没有利用主基因+多基因模型分析抗旱性状的报道。本研究以抗旱品种定亚 17 号为母本,抗旱敏感品种天亚 3号为父本,采用一粒传法构建RIL群体,利用数量性状主基因+多基因混合遗传模型分析方法,研究胡麻苗期抗旱相关性状的遗传机制,明确胡麻苗期相关性状抗旱的遗传因素和环境变异,为胡麻抗旱育种提供理论指导。
1 材料与方法
1.1 试验材料
以抗旱品种定亚 17 号为母本(P1)、抗旱敏感品种天亚 3号为父本(P2),于2012年12月份在云南元谋杂交,采用一粒传法在兰州和元谋两地连续自交7代构建了F7代139个株系的RIL群体。试验用RIL群体和两个亲本群体进行遗传分析。
1.2 试验方法
试验在甘肃省农业科学院智能温室进行抗旱性鉴定。试验设置干旱胁迫和对照(正常灌水)2个处理,出苗后进行干旱胁迫,正常灌水CK(田间最大持水量的80%),干旱胁迫(田间最大持水量的50%)。每个处理3次重复,小区按2 m行长、20 cm行距、3行区种植,每行播种300粒。土壤水分数据采用EM50(Decagon Device. USA)数据采集器(采集时间间隔为1h)进行采集,埋在不同处理0~30 cm深的土壤中实时监测土壤中的水分动态,根据水分的实际情况确定灌水量。田间最大持水量用环刀法测定。
4月20号播种,6月初取样。每小区取样10 株,挖出根后用水冲洗干净,带回实验室,用直尺测定主根长和株高;用万分之一天平测定根茎叶鲜重。然后于105℃烘干箱内杀青1h,再在80℃恒温下烘6~8h至恒重,测定其干重。共考察根长(Root length,RL)、株高(Plant height,PH)、叶鲜重(Fresh leaf weight,FLW)、叶干重(Leaf dry weight,LDW)、根鲜重(Fresh root weight,FRW)、根干重(Root dry weight,RDW)、茎鲜重(Fresh shoot weight,FSW)、茎干重(Fresh shoot weight,FSW)8个性状,计算抗旱系数。
DC=Xi/CKi
式中,Xi、CKi分别表示干旱、对照处理条件下性状测定值。
1.3 数据分析
采用盖钧镒、章元明[14,15]等提出的数量性状主基因+多基因混合遗传分离分析方法,对两个亲本群体和RIL群体的8个苗期性状的抗旱系数进行遗传模型分析,运行SEA-G3DH分析软件进行数据处理。首先,计算不同性状遗传模型的极大似然函数值(Max-likelihood-values,MLV)和AIC(Akaike's information criterion)值。根据AIC 筛选方法,首先选定AIC 值最小的3个模型作为备选模型,通过对备选模型进行适合性检验,有5 个统计量:U12、U22、U23(均匀性检验)、nW2(Smirnov 检验)和Dn(Kolmogorov 检验),然后选择的最优模型为AIC值比较小且统计量达到显著水平个数最少的模型。在此基础上,采用最小二乘法估计遗传模型的一阶、二阶遗传参数,包括主基因加性、显性、上位性效应,多基因的加性和显性效应,主基因和多基因方差及遗传率等[22]。
2 结果与分析
2.1 不同群体相关性状抗旱系数的分布
两个亲本群体及RIL群体8个性状抗旱系数平均值的统计值和正态性检验值列于表1。从变异系数看,根鲜重、茎鲜重、根干重和茎干重的变异系数较大(42.54~48.39%),株高和根长的变异系数相对较小(18.37%和16.93%)。8个性状抗旱系数的平均值均呈连续性分布,其分布特征函数偏度和峰度绝对值都小于1,其中株高呈左偏离正态分布,其它性状呈右偏离正态分布;根鲜重呈平顶峰分布,其它性状呈尖顶峰分布(图1)。株高,根长和茎干重的正态性检验P0.05值均大于0.05,其它性状的正态性检验P0.05值都小于0.05。表明这些性状分布近似呈正态分布,可能受主效基因和微效多基因效应共同控制。
2.2 不同性状遗传模型选择及适合性检验
利用植物数量性状主基因+多基因混合遗传模型,对RIL群体和两个亲本群体的8个性状的抗旱系数进行模型分析,运行SEA-G3DH分析软件,得到不同性状的不同模型极大对数似然值(MLV)和AIC 值(表2)。
根据AIC 值最小原则进行模型初选,从中选择AIC值最小的模型,同时选择与其接近的两个模型作为备选模型。从表2可以看出,株高AIC值最小的模型为2MG-Duplicate(-245.609),与其接近的模型为3MG-CEA(-270.594)和2MG-Inhibiting(-281.128);分别对这3个模型的3个群体进行适合性检验(表4),包括均匀性检验(U12、U22、U32),Smirnov检验(nW2)和Kolmogorov 检验(Dn),从3个模型中选择统计量达到显著水平个数最少的模型为最优模型。株高3个备选模型的3个群体共15个统计量中达到显著水平的统计量个数分别是13、11、11,因此株高的最适遗传模型为2MG-Inhibiting,受2 对抑制性主基因+加性多基因模型控制(表3)。同样,根鲜重、茎鲜重、根干重、茎干重的最适遗传模型为2MG-Inhibiting。根长、叶干重的最适遗传模型为3MG-CEA,受3对独立主基因+加性多基因模型控制;叶鲜重的最适遗传模型为4MG-CEA,受4对独立主基因控制,没有多基因效应。胡麻苗期8个抗旱相关性状有5个受2 对主基因+多基因遗传控制,2个受3对主基因+多基因遗传控制,1个受4对独立主基因控制。有5个性状的主基因遗传效应为抑制性作用,3个性状为独立主基因;7个性状的多基因遗传效应为加性作用。
2.3 遗传参数估计
采用最小二乘法估计8个性状抗旱系数最适遗传模型的一阶、二阶遗传参数(表4)。株高、根鲜重、茎鲜重、根干重、茎干重、根长、叶干重7个性状受主基因+多基因遗传控制,叶鲜重只有主基因效应,没有多基因效应。株高、根鲜重、茎鲜重、根干重、茎干重的2对主基因为抑制效应,多基因效应为加性效应,只有根鲜重的加性效应值是负向,其它的都为正向;这5个性状的主基因遗传率分别是19.61%、41.67%、37.58%、9.59%、50.96%。根长、叶干重的3对独立主基因效应完全相等,而且效应值全是负向;多基因为加性效应;这两个性状的主基因遗传率分别是41.97%和34.73%。叶鲜重的4对独立主基因效应完全相等,效应值全是负向;没有多基因效应,这个性状的主基因遗传率为46.35%。
表1不同性状抗旱系数的统计值和正态性检验
Table 1 Statistic analysis and normal distribution test of different traits drought resistance coefficient
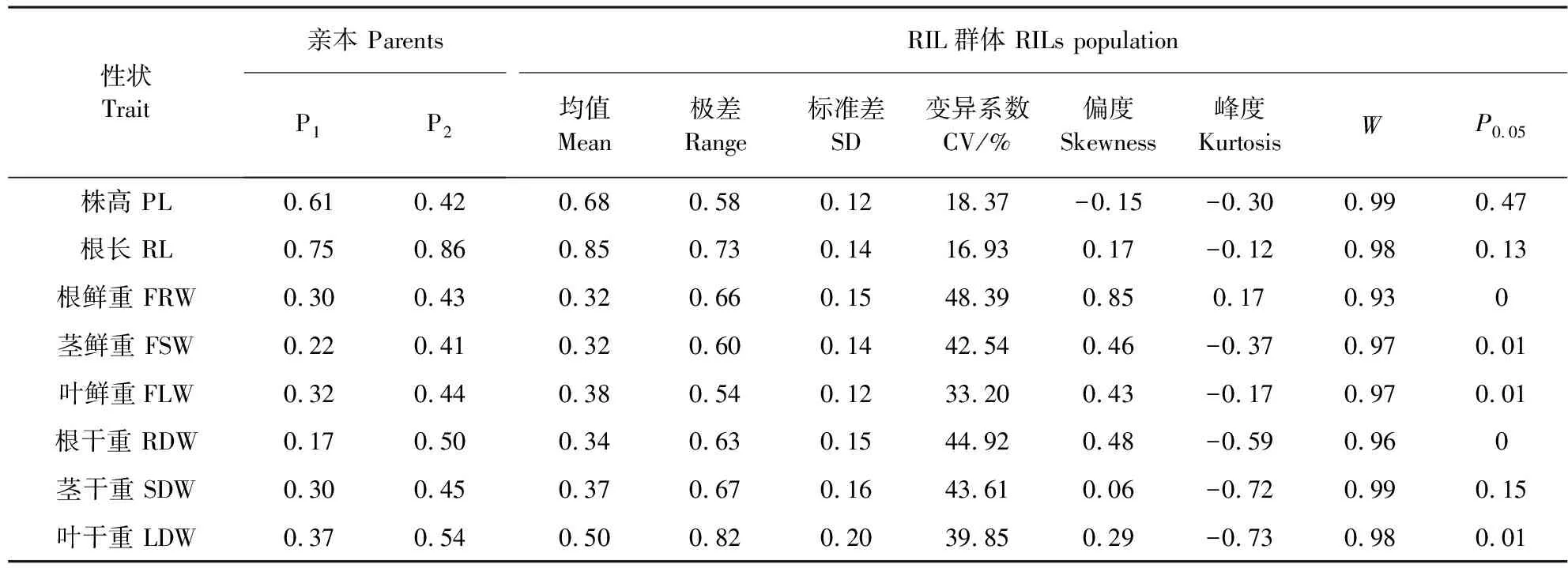
性状Trait亲本 ParentsP1P2RIL群体 RILs population均值Mean极差Range标准差SD变异系数CV/%偏度Skewness峰度KurtosisWP0.05株高 PL0.610.420.680.580.1218.37-0.15-0.300.990.47根长 RL0.750.860.850.730.1416.930.17-0.120.980.13根鲜重 FRW0.300.430.320.660.1548.390.850.170.930茎鲜重 FSW0.220.410.320.600.1442.540.46-0.370.970.01叶鲜重FLW0.320.440.380.540.1233.200.43-0.170.970.01根干重 RDW0.170.500.340.630.1544.920.48-0.590.960茎干重 SDW0.300.450.370.670.1643.610.06-0.720.990.15叶干重 LDW0.370.540.500.820.2039.850.29-0.730.980.01

图1 不同性状抗旱系数次数分布图Fig.1 Frequency distribution of different traits drought resistance coefficient
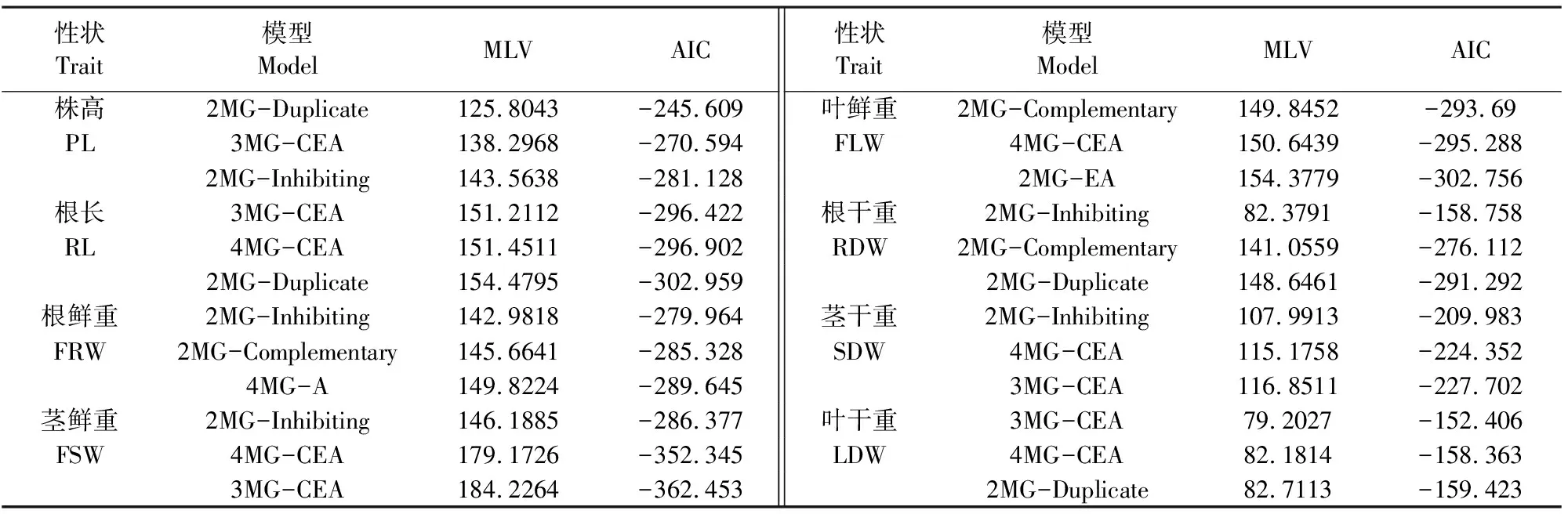
性状Trait模型ModelMLVAIC性状Trait模型ModelMLVAIC株高2MG-Duplicate125.8043-245.609叶鲜重2MG-Complementary149.8452-293.69PL3MG-CEA138.2968-270.594FLW4MG-CEA150.6439-295.2882MG-Inhibiting143.5638-281.1282MG-EA154.3779-302.756根长3MG-CEA151.2112-296.422根干重2MG-Inhibiting82.3791-158.758RL4MG-CEA151.4511-296.902RDW2MG-Complementary141.0559-276.1122MG-Duplicate154.4795-302.9592MG-Duplicate148.6461-291.292根鲜重2MG-Inhibiting142.9818-279.964茎干重2MG-Inhibiting107.9913-209.983FRW2MG-Complementary145.6641-285.328SDW4MG-CEA115.1758-224.3524MG-A149.8224-289.6453MG-CEA116.8511-227.702茎鲜重2MG-Inhibiting146.1885-286.377叶干重3MG-CEA79.2027-152.406FSW4MG-CEA179.1726-352.345LDW4MG-CEA82.1814-158.3633MG-CEA184.2264-362.4532MG-Duplicate82.7113-159.423
表3不同性状抗旱系数遗传模型的适合性检验
Table 3 Test for fitness of genetic models on different traits drought resistance coefficient

性状Trait模型Model世代GenerationU12(p)U22(p)U32(p)nW22(p)Dn(p)株高2MG-InhibitingP174.71(0.00)44.98(0.00)44.21(0.00)8.36(0.01)0.79(0.00)PLP25.99(0.01)1.35(0.24)23.28(0.00)1.71(0.00)0.21(0.054)RILs9.67(0.00)6.39(0.01)3.74(0.053)1.30(0.00)0.02(1.00)根长3MG-CEAP10.80(0.37)5.77(0.02)37.72(0.00)1.60(0.00)0.37(0.00)RLP214.62(0.00)21.44(0.00)13.78(0.00)2.82(0.00)0.50(0.00)RILs0.89(0.34)2.08(0.15)4.48(0.03)0.20(0.26)0.01(1.00)根鲜重2MG-InhibitingP117.07(0.00)23.06(0.00)10.28(0.00)2.93(0.00)0.52(0.00)FRWP223.11(0.00)16.98(0.00)4.56(0.03)3.45(0.00)0.14(0.41)RILs2.22(0.14)1.06(0.30)2.76(0.10)0.61(0.02)0.01(1.00)茎鲜重2MG-InhibitingP123.32(0.00)27.08(0.00)4.46(0.03)3.49(0.00)0.51(0.00)FSWP256.29(0.00)66.09(0.00)11.97(0.00)6.65(0.01)0.06(1.00)RILs1.88(0.17)0.93(0.33)2.10(0.15)0.30(0.15)0.01(1.00)叶鲜重4MG-CEAP10.79(0.37)0.36(0.55)33.96(0.00)1.32(0.00)0.28(0.00)FLWP22.42(0.12)8.73(0.00)33.53(0.00)1.67(0.00)0.40(0.00)RILs1.75(0.19)1.25(0.26)0.41(0.52)0.37(0.09)0.01(1.00)根干重2MG-InhibitingP150.73(0.00)40.06(0.00)5.15(0.02)6.56(0.01)0.74(0.00)RDWP262.03(0.00)75.65(0.00)18.38(0.00)7.52(0.01)0.07(0.98)RILs1.40(0.24)1.54(0.22)0.15(0.70)0.29(0.15)0.00(1.00)茎干重2MG-InhibitingP18.00(0.00)15.69(0.00)23.93(0.00)2.24(0.00)0.48(0.00)SDWP244.28(0.00)45.35(0.00)1.35(0.24)5.90(0.00)0.11(0.74)RILs1.75(0.19)0.81(0.37)2.31(0.13)0.22(0.24)0.00(1.000)叶干重3MG-CEAP110.89(0.00)19.08(0.00)22.02(0.00)2.89(0.00)0.53(0.00)LDWP232.15(0.00)32.98(0.00)1.02(0.31)4.90(0.00)0.65(0.00)RILs6.64(0.01)10.63(0.00)9.40(0.00)0.80(0.01)0.01(1.00)
表4不同性状抗旱系数遗传模式的遗传参数估计值
Table 4 Estimates of genetic parameters from different traits drought resistance coefficient
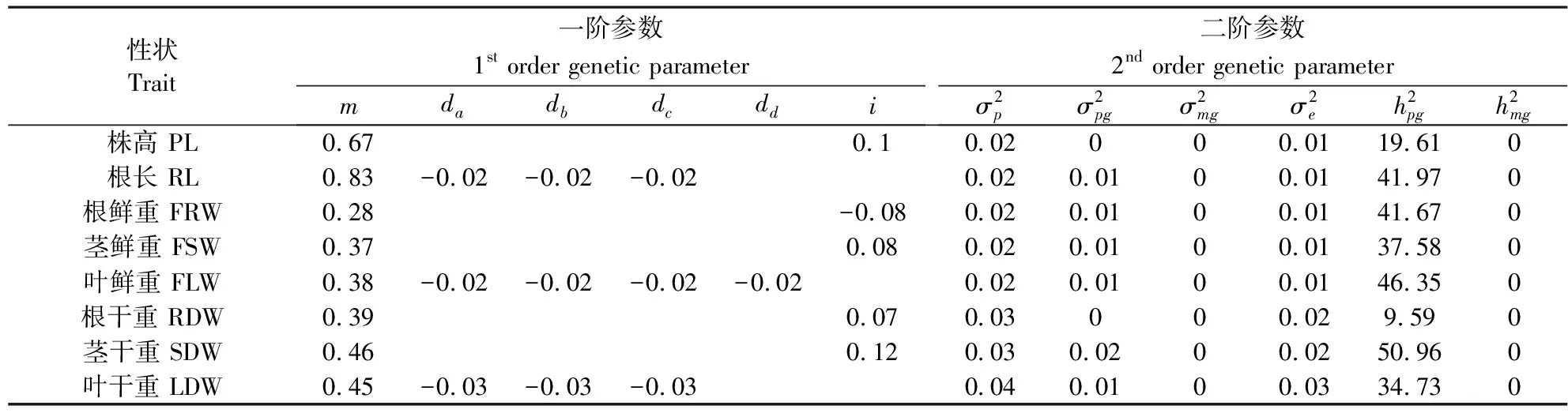
性状Trait一阶参数1st order genetic parametermdadbdcddi二阶参数2nd order genetic parameterσ2pσ2pgσ2mgσ2eh2pgh2mg株高 PL0.670.10.02000.0119.610根长 RL0.83-0.02-0.02-0.020.020.0100.0141.970根鲜重 FRW0.28-0.080.020.0100.0141.670茎鲜重 FSW0.370.080.020.0100.0137.580叶鲜重 FLW0.38-0.02-0.02-0.02-0.020.020.0100.0146.350根干重 RDW0.390.070.03000.029.590茎干重 SDW0.460.120.030.0200.0250.960叶干重 LDW0.45-0.03-0.03-0.030.040.0100.0334.730

3 讨 论
相关研究表明,胡麻数量性状的遗传受加性和非加性(显性、上位性)基因效应共同控制[17,18]。本研究结果表明,株高、根鲜重、茎鲜重、根干重、茎干重受2对抑制性主基因+加性多基因控制,根长、叶干重受3对独立主基因+加性多基因控制,叶鲜重受4对独立主基因控制,没有多基因效应。从主基因的遗传效应大小及方向看,不同性状存在差异比较大。从主基因遗传效率来看, 8个性状主基因遗传率在9.59%~50.96%之间,株高和根干重遗传率比较低,说明环境对这两个性状影响比较大。胡标林等对东乡野生稻BILs群体苗期抗旱性遗传分析显示受2 对独立主基因+多基因控制[23],刘莹等对大豆耐旱种质相关根系性状的遗传分析显示,耐旱相关根系性状主要由2对主基因和加性多基因控制,与本研究结果相近[24]。闫艳等研究认为,小麦苗期根长、根鲜重、茎干重、根干重等性状在干旱条件下受1对加性显性主基因控制,茎鲜重受2对加性显性主基因控制,苗高无主基因,与本研究结果不同[25]。朱猛等分析表明,干旱胁迫下玉米苗期主根长和根鲜重的遗传模型为1对加性-显性主基因+加性-显性-上位性多基因控制[26]。说明不同作物抗旱性数量性状比较复杂,不同的试验结果可能与试验材料、试验环境、选择的指标等差异有关。根的生长是作物在干旱胁迫下所需观察的重要性状,苗期根系性状应该作为一项重要的参考指标,但在大田进行根性状的调查误差比较大[27]。周晓果等对小麦 DH 群体苗期根系性状的研究表明,有3个加性效应QTL和3对上位性效应QTL控制最大根长,2个加性效应QTL和2对上位性效应QTL控制根鲜重,2个加性效应QTL和3对上位性效应QTL影响根干重。控制这些性状的主基因数目和本文研究结果相同,但主基因效应不同[28]。翟荣荣等在不同水分条件下水稻苗期根系性状的 QTL 分析显示,在干旱胁迫条件下检测到 14个QTL,影响根长的QTL有2个,影响根鲜重的QTL有3个[29]。Tuberosa 等[30]在玉米抗旱研究中和周蓉等[31]在大豆抗旱性研究中都发现根植株抗旱性与产量性状存在连锁关系。水稻上研究表明根系性状对植株抗旱性[32,33],地上部生长和产量形成起着重要作用[34-36],说明根系性状与作物抗旱性密切相关。本研究显示,根长、根鲜重、叶鲜重和茎干重的主基因遗传率比较高,可以作为胡麻苗期抗旱的鉴定指标。根鲜重、茎鲜重、根干重和茎干重的变异系数最大,说明4个性状存在较广泛的变异,并且存在超亲优势,选择优势株系进行新品种选育的潜力比较大。
4 小 结
胡麻RIL 群体8个性状的抗旱系数存在广泛变异,部分性状表现超亲分离现象,其分布近似为正态分布。通过主基因+多基因遗传模型分析表明胡麻RIL群体苗期抗旱性的8个性状中有7个为主基因+多基因混合遗传模型,株高、根鲜重、茎鲜重、根干重、茎干重受2对抑制性主基因+加性多基因控制,根长、叶干重受3对独立主基因+加性多基因控制,叶鲜重受4对独立主基因控制。从主基因的遗传效应大小及方向看,不同性状存在差异比较大。总体来看,抑制性和独立基因主基因,加性多基因在抗旱相关性状的遗传中起重要作用。从主基因遗传效率来看,根长、根鲜重、叶鲜重和茎干重可以作为胡麻苗期抗旱鉴定指标。
