油用牡丹凤丹光合作用对光照度及胞间CO2浓度的季节响应
2018-10-16杨玉珍王国霞张志浩
杨玉珍,王国霞,张志浩
(1.郑州师范学院生命科学学院,河南郑州450044;2.郑州大学生命科学学院,河南郑州450001)
牡丹(Paeonia suffruticosa Andr.)原产于中国,为芍药科(Paeoniaceae)芍药属(Paeonia)多年生木本落叶小灌木,在我国已有1 600多年的栽培历史[1],素有“国色天香”“花中之王”的美称。油用牡丹为近几年新兴的一种优质木本油料作物,籽油是其主要产品之一,具有极高的营养价值。2011年3月,牡丹籽油被卫生部正式批准为新资源食品[2]。目前,油用牡丹在全国已大面积推广种植,以凤丹(Paeonia ostii)和紫斑(Paeonia rockii)为主要栽培种。其中,凤丹牡丹具有耐湿热、株型高大、结实率高、适应性强的特性,分布于全国大部分地区,特别是在河南、安徽、山东等省份已有大面积种植;紫斑牡丹由于耐寒抗旱性很强,主要分布在四川、甘肃、陕西等西部地区[3]。
在油用牡丹的引种栽培中,光和CO2是重要的生态因子,直接影响着光合作用及最终产量。在关于光合作用的研究中,光合作用的响应是研究植物生理和植物生态学的重要途径。由光响应曲线可以确定植物的光补偿点、光饱和点、表观量子效率及最大净光合速率;由CO2响应曲线可以确定植物的CO2补偿点、饱和点、RuBP羧化作用以及最大净光合作用。这些指标对了解植物的生理特性具有指导作用,因此,确定植物光合作用对主要生态因子如光照和细胞间CO2浓度在不同季节里的响应,对于研究植物的光合特性、指导农业生产方面具有重要意义[4]。目前,植物光合作用的光响应模型有很多,各有优劣,有直角双曲线模型[5]、非直角双曲线模型[6]、直角双曲线的修正模型[7]等,而在对油用牡丹的光响应研究中应用最多的是非直角双曲线模型[7-8],本试验采用该模型拟合。CO2响应模型主要有生化模型[9]、直角双曲线模型[10]、直角双曲线修正模型[11]等,本研究首次将直角双曲线模型应用于油用牡丹的CO2响应。本研究以油用牡丹主要优良栽培品种凤丹为研究对象,通过比较不同季节条件下其光合作用对光合有效辐射和胞间CO2浓度响应的差异,旨在为油用牡丹在河南地区的大面积推广栽培以及生产管理提供理论指导。
1 材料与方法
1.1 材料
供试材料为5年生凤丹白品种(Paeonia ostii T.Hong et J.X.Zhang‘Phoenix White’),种植于郑州师范学院黄河滩实习实训基地。试验地处北温带大陆性季风气候区,34°51'N、113°38'E,海拔110 m,冷暖适中、四季分明,春季干旱少雨,夏季炎热多雨,秋季晴朗日照长,冬季寒冷少雪,年平均气温在14~14.3℃之间,平均降水量640.9 mm,无霜期220 d,全年日照时间约2 400 h。植物材料大田种植,水肥条件良好,常规管理。周边无高大遮挡物,处全光照环境。原位随机选取测定长势基本一致、无病虫害的健康植株,顶端往下第3张或第4张向阳功能叶,尽量保持叶片正常生长角度不变。
1.2 方法
1.2.1 光响应曲线的测定 分别于2016年5月下旬(春季)和2016年7月下旬(夏季)连续晴天运用LI-6400XT便携式光合测量系统(LI-COR,USA)进行瞬时光合速率的测定。测定时间为09:00—11:00。试验重复3次,每次选择1个典型植株。以CO2注入系统为气源,浓度设定为接近于大气CO2浓度400μmol/mol。测定前打开2 000μmol/(m2·s)的6400-02B型LED灯进行光诱导。光照度设置为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、80、60、40、20、0 μmol/(m2·s),共 16 个梯度值。程序设定完成后,仪器自动记录数据。数据导出后,依据非直角双曲线模型进行光响应曲线拟合[6],拟合数据取3次重复的平均值,拟合公式如下:

式中:Pn为净光合速率,Pn,max为最大净光合速率,Q为光合有效辐射,o为表观量子效率(apparent quantum yield,AQY),k为拟合曲线曲角,Rd为暗呼吸速率。将光照度在200μmol/(m2·s)以下阶段以直线回归作补充,与非直角双曲线模型下计算出的 Pn,max及对应的光照度,即光饱和点(light saturation point,LSP)相接,与x轴的交点即为光补偿点(light compensation point,LCP),直线斜率即为 AQY。
1.2.2 CO2响应曲线的测定 分别于2016年5月下旬(春季)和2016年7月下旬(夏季)连续晴天运用LI-6400XT便携式光合测量系统测定凤丹CO2响应曲线。测定时间在09:00—11:00,试验重复3次,每次选择1个典型植株。光照度设定在1 000μmol/(m2·s),由6400-02B型红蓝光源提供。叶室温度设定为25℃。CO2浓度设定为50、100、200、300、400、600、800、1 000、1 200、1 500、1 800、2 000 μmol/mol共计12个梯度值,每个梯度下最小等待时间为120 s,最大等待时间为300 s。程序设定完成后,仪器自动记录数据。数据导出后,利用直角双曲线[10]拟合细胞间CO2浓度Ci与净光合速率Pn,拟合数据取3次重复的平均值,拟合公式如下:

式中:Ci为胞间CO2浓度,CE为RuBP羧化效率,Rp为光呼吸速率(由于光下暗呼吸速率很小,可近似等于光下叶片向外界释放CO2浓度的速率)。将低CO2浓度在200μmol/mol以下阶段以直线回归作补充,与直角双曲线模型下计算出的Pn,max及对应的 CO2浓度即 CO2饱和点(CO2Saturation Point,CSP)相接,则与x轴的交点即为CO2补偿点(CO2compensation point,CCP),截距为 Rp,CO2浓度 200 μmol/(m2·s)以下阶段直线斜率即为RuBP羧化效率CE。
1.3 数据处理
试验数据采用Excel 2003及SPSS 20.0软件进行分析,Origin 8.0 作图。
2 结果与分析
2.1 凤丹净光合速率对光合有效辐射的季节响应
2.1.1 不同季节光响应初始斜率比较 AQY作为叶片光能利用效率重要参数之一,反映了植物在进行光合作用时对弱光的利用效率,可用光响应曲线的初始斜率[0~200μmol/(m2·s)光照度范围]计算得到。一般情况下,植物的 AQY 值在0.03~0.07之间[12]。由表1可以看出,在春季,大田条件下凤丹光响应曲线的初始斜率略小于一般值,而夏季数值在正常范围,表明凤丹在夏季较春季对弱光的利用效率高。

表1 不同季节凤丹光响应曲线的初始斜率及相应的回归方程
2.1.2 不同季节光响应曲线及相关参数比较 光饱和点和补偿点能反映植物对强光和弱光的利用能力,是反映植物光响应的重要指标[13]。由图1可知,凤丹叶片在不同季节的光响应曲线变化趋势相同。在光合有效辐射为0μmol/(m2·s)时,“凤丹”的净光合速率均为负值,在0~200μmol/(m2·s)光照度范围内,随着光合有效辐射(Photosynthetically ctive radiation,PAR)的增加,Pn近乎直线增长,直到接近 Pnmax,此时对应的PAR值为LSP。通过对0~200μmol/(m2·s)光照度范围内曲线进行线性回归及用非直角双曲线模型对>200μmol/(m2·s)光照度范围曲线进行拟合(表2),经计算发现,在春季和夏季的 LSP分别为 908.547、1 355.527μmol/(m2·s)。净光合速率为 0 μmol/(m2·s)时,光合有效辐射为 LCP,在春季和夏季分别为36.087、54.083μmol/(m2·s),说明凤丹在春季对弱光的利用能力强于夏季。最大净光合速率Pn,max为植物在饱和光照度下的净光合速率,是叶片的最大净光合速率,能够反映叶片的光合效率和光合潜能。该值在2个季节具有明显差别,分别为24.421、41.059 μmol/(m2·s)。暗呼吸速率 Rd可以反映呼吸速率,在春季和夏季分别为0.968、1.711 μmol/(m2·s)。


表2 不同季节凤丹光合-光照度响应主要参数拟合值
2.2 凤丹净光合速率对细胞间CO2浓度的季节响应
2.2.1 不同季节CO2响应初始斜率比较 RuBP羧化效率CE是指低CO2浓度(0~200μmol/mol)的净光合速率与胞间CO2浓度拟合的一次方程的斜率,是衡量植物对叶片中CO2利用效率的重要指标[14]。CE值越大,说明在低CO2浓度条件下能够更高效地利用CO2进行光合作用。由表3可知,凤丹在春季CE值显著高于夏季,说明在春季对低CO2浓度的利用高于夏季。
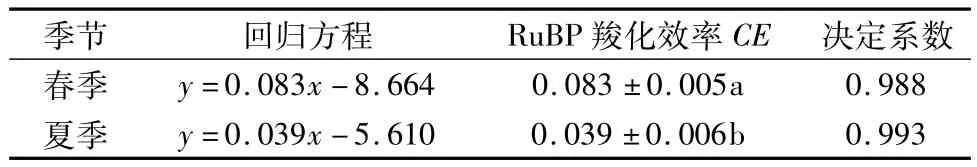
表3 不同季节凤丹CO2响应曲线的初始斜率及相应的回归方程
2.2.2 不同季节CO2响应曲线比较 CO2作为光合作用的底物,其浓度在一定范围内直接影响着光合作用的进程。图2显示,随着Ci值的增大,净光合速率增大,且春季增幅明显大于夏季。在低Ci浓度(小于200μmol/mol)时,Pn上升速度较快,一般认为,此时1,5-二磷酸核酮糖羧化酶(rubsico)对Pn的限制起主要作用。此后,随着Ci浓度的增大,Pn增幅减小,直至接近饱和状态,此阶段的1,5-二磷酸核酮糖羧化酶再生速率是制约Pn的主要原因。在不同季节分别达到相应的CO2饱和点(1 152.506、1 425.059 μmol/mol)时,春季的最大净光合速率Pn,max显著高于夏季。然而,夏季相对较低的CO2补偿点说明在夏季凤丹对低CO2浓度的利用效率高于春季,可见凤丹在夏季具有较宽的CO2适应性。与此同时,在春季还伴随着较高的光呼吸速率(表4)。


表4 不同季节凤丹光合-C i响应主要参数拟合值
3 结论与讨论
光合作用为地球上的各种生物提供了赖以生存和发展的食物、空气和能量,是一个十分复杂的生理过程。不同植物叶片的光合特性既受自身遗传因素影响,也与光照等环境因子有关。掌握凤丹在不同季节条件下的生长规律,有助于提高产量,具有重要的理论和指导意义。
3.1 凤丹光合作用对光的季节响应
光饱和条件下的净光合速率有时被称为光合能力,或光合潜力[10]。光饱和点和补偿点分别反映植物对强光和弱光的利用情况,同时也与植物的生长阶段和适应对策以及不同季节的气候条件密切相关。本试验中夏季凤丹具有较高的光饱和点,说明其对强光利用能力优于春季,而较高的光补偿点伴有较高的暗呼吸速率也说明凤丹对弱光的利用能力低于春季。光合机构每吸收1个光量子固定的CO2或释放的O2分子数被称为光合碳同化的量子效率。AQY不考虑叶片光透射和反射(15%左右),而是按照入射的光量子数计算得到的量子效率[14]。植物的表观量子效率越高,其对光能的利用能力就越强。本研究中夏季凤丹AQY值大于春季,说明其在夏季对光能的利用能力较强。光补偿点在农业生产实践中有很大的意义。在间作和套作中选择搭配的作物种类,林带树种的配置等都与光补偿点有关。栽培过密或水肥过多,容易造成徒长,使得封行提前,作物中下部叶片接受的光照匮乏,低于光补偿点,叶片不但不能制造有机物,反而会消耗自身养分,导致作物减产。另外,毕玉伟等以盆栽条件下的凤丹为研究对象得出的光饱和点和补偿点范围均大于本研究结果,推测盆栽和田间气候因子的差异可能是造成这种不符情况的主要原因,也可能与Pn-PAR数据拟合的方法[15]有关。
3.2 凤丹光合作用对胞间CO2浓度的季节响应
普通空气中的CO2浓度偏低,制约了光合作用。CO2是光合作用的原料,是RuBP羧化酶的主要底物,其补偿点和饱和点是植物是否具有潜在高光合效率的重要指标。大田条件下凤丹在春季和夏季的 CCP分别为 138.184、54.256μmol/mol,属一般C3植物范畴,说明在夏季凤丹对低浓度CO2的利用能力优于春季;春季的羧化效率高于夏季,表明凤丹在春季对低CO2浓度适应性更强。在达到远高于大气CO2浓度值400μmol/moL左右的CSP时,凤丹的最大Pn,max可分别达 86.603、54.256 μmol/(m2·s),远大于同时期自然条件下的最高净光合速率,说明凤丹还具有巨大的光合潜力。在一定的CO2浓度范围内,适当增施CO2肥会极大促进油用牡丹产量。
