轮状病毒NSP5结构与功能的研究进展
2018-10-15刘洋林晓晨李鸿钧
刘洋, 林晓晨, 李鸿钧
(1. 昆明医科大学, 昆明 650500; 2. 中国医学科学院 北京协和医学院 医学生物学研究所, 云南省重大传染病疫苗研发重点实验室, 昆明 650118)
在全球范围内经研究证实,轮状病毒为5岁以下儿童发生严重感染性腹泻的常见病毒,因此由轮状病毒感染引发的疾病受到关注。
轮状病毒是由11个双链RNA片段通过有序的拆卸和组装三层结构的二十面体来进行管理和复制(见图1)。基因片段1负责结构蛋白VP1、基因片段2负责结构蛋白VP2、基因片段3负责结构蛋白VP3、基因片段4负责结构蛋白VP4、基因片段5负责非结构蛋白NSP1、基因片段6负责结构蛋白VP6、基因片段7负责非结构蛋白NSP2、基因片段8负责非结构蛋白NSP3、基因片段9负责结构蛋白VP7、基因片段10负责非结构蛋白NSP4、基因片段11负责非结构蛋白NSP5和非结构蛋白NSP6[1-2]。VP1蛋白是在组装病毒RNA时必需的聚合酶;VP2蛋白是将VP1及VP3蛋白装配其内部的基质蛋白;VP4蛋白是由VP5蛋白和VP8蛋白组成的结合蛋白,以及形成双层颗粒结构,进而可以合成病毒mRNA的组抗原蛋白VP6;VP7是同VP4共同组成最外层衣壳结构的外壳糖蛋白[3]。NSP1的作用方式是通过对干扰素调节因子(IRF)的影响,进而发挥作用;通过判定NSP2在机体内的抗体水平,进而判断是否再度感染轮状病毒;NSP3的作用需进一步研究;NSP4通过对细胞钙稳态的影响作用,进而在细胞形态、致病机制中都发挥重要作用;NSP5在病毒复制和转录中是不可缺少的重要因素。
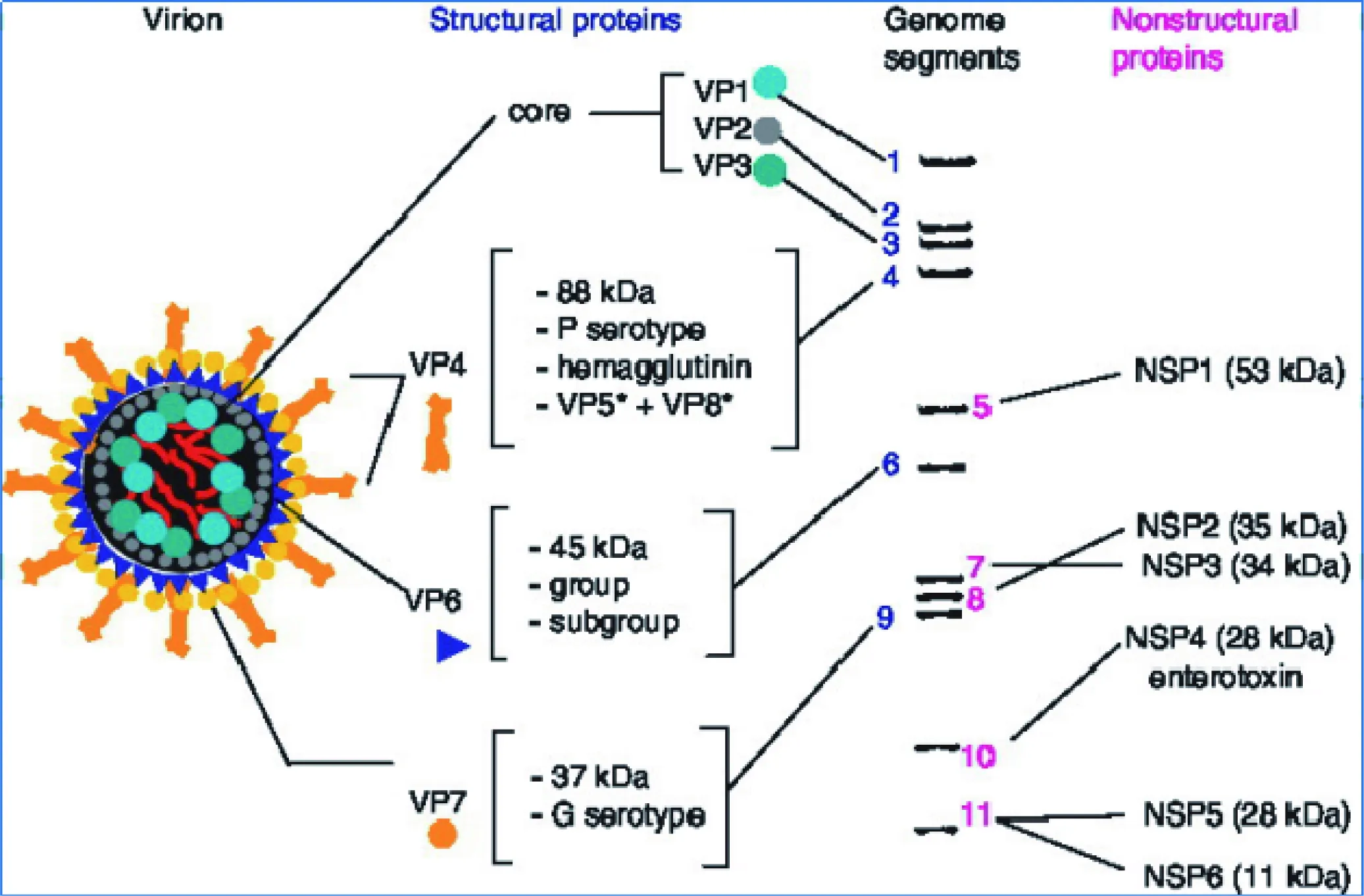
图1 轮状病毒结构[4]
1 轮状病毒NSP5的结构相关研究
Martin等[5]通过多角度激光散射、沉降速度和平衡沉降实验揭示了NSP5具有两个低聚区域,第一个区域残基103~146,参与NSP5二聚化;而第二区域残基189~198,负责NSP5十聚体,通过分析十聚体区域主要是螺旋的,而二聚区域涉及β-折叠,进一步研究显示NSP5包含两α-螺旋,无序的N-末端和C-末端是主要由β-折叠层组成。研究中还发现,NSP5片段和NSP2在未感染细胞中的共表达NSP5十聚体区域是形成类病毒所必需的结构。然而,在体外,NSP5的区域部分抑制NSP2-NSP5相互作用,研究中的NSP5模型表明该空间位阻阻止NSP2与所有NSP5的结合。NSP5是一种富含苏氨酸且丝氨酸被高度磷酸化的高酸性蛋白。NSP5还是一个可通过不同翻译后修饰,表达为26~35 ku不同分子质量的糖基化蛋白。轮状病毒NSP5结构尚不能完全确定,研究发现在编码内衣壳蛋白VP6和非结构蛋白NSP2,NSP4和NSP5的保守的基因中,存在一定程度的高遗传性[6-8]。
Blackhall等[9]研究表明轮状病毒非结构蛋白NSP5由RNA片段11中的基因编码。NSP5在COS-7细胞中表达时,在不受其他病毒蛋白干扰条件下,其均匀分布在细胞质中。在这些条件下,26 ku多肽占优势。在蛋白磷酸酶抑制剂存在下,高度磷酸化形成28 ku和32 ku至35 ku的多肽。此外,完全磷酸化的蛋白质在转染的细胞中具有均匀的细胞质分布。在轮状病毒SA11感染细胞中,感染后2 h可检测到NSP5合成。然而,新形成的26 ku转化为28~35 ku的形式需要大约2 h左右。从转染COS-7细胞中提取的NSP5体外检测,是有自身磷酸化的功能,表明其不需要其他病毒蛋白参与进行磷酸化。用星形孢菌素(广泛的蛋白激酶抑制剂)处理表达NSP5的细胞,对病毒多肽的磷酸化影响有限,说明星孢菌素没有抑制NSP5体外自磷酸化。
轮状病毒NSP5蛋白与病毒样结构(VLS)的形成有关。在Sen等[10]研究报告中,NSP5的C末端氨基酸是形成VLS所必需的,是与NSP5的不溶性和高度磷酸化有关。通过NSP5的C末端计算机建模和分析,显示存在跨越C末端残基的两亲性α-螺旋。通过研究发现伴随着螺旋被破坏,NSP5在VLS中定位也消失。这些研究确定了调节VLS形成的NSP5C末端内的功能决定簇,并提供了在轮状病毒复制过程中抑制NSP5定向VLS功能的靶标。
2 轮状病毒NSP5的相关功能研究
轮状病毒(RV)复制发生在称为病毒基质的细胞质内含物中,轮状病毒的复制需要非结构蛋白NSP2和非结构蛋白NSP5的相互作用才能进行[11]。NSP5和NSP2之间的相互作用推动了病毒载体的形成,以及轮状病毒感染细胞中基因组复制和包装的位点。富含丝氨酸-苏氨酸的NSP5在复制循环期间在低磷酸化和高磷酸化异构体之间转变。
Jiang等[12]为揭示NSP2如何与NSP5和RNA的相互作用,他们单独进行NSP2、NSP5和ssRNA复合物的单粒子冷冻电子显微镜(cryo-EM)分析。研究表明,NSP2的结构与晶体八聚体相同,并且NSP5 66-188和ssRNA都与八聚体中带正电荷的残基凹槽结合。
轮状病毒是在病毒感染细胞的细胞质中形成的。Eichwald等[13]揭示了非结构蛋白NSP2和NSP5与其他病毒蛋白一起定位在病毒中,包括VP1、VP3和VP2。通过酵母双杂交系统和体内结合/免疫沉淀这两种不同的测定法,测定研究NSP2与一系列NSP5突变体的相互作用,表明NSP5 N-末端区域及C-末端部分区域需要绑定到NSP2才能发挥作用。
Martin等[14]的研究表明,非结构蛋白NSP5还是可以结合一个[2Fe-2S]簇的病毒金属蛋白。研究发现,VP1在病毒复制过程中带有[2Fe-2S]簇的非结构蛋白NSP5是辅助因子,结合[2Fe-2S]簇的非结构蛋白NSP5在与单链RNA结合时,其亲和力也得到了增强。
Campagna等[15-16]通过靶向基因组区段11mRNA的siRNA干扰两种不同的菌株,以特异性方式阻断了NSP5的翻译,发现在病毒的复制周期中都产生了很强的影响作用,包括:病毒的形成受到了抑制,结构蛋白和非结构蛋白表达合成减少,病毒基因组dsRNA的合成减少及传染性的子代病毒合成减少,说明在病毒的复制和组装中NSP5是必需的。还发现,通过点突变使NSP5的SUMO修饰位点发生改变后,VLS形成受阻,病毒因此无法完成复制。Taraporewala等[17]研究表明,轮状病毒感染细胞,在体内复制需要一个稳定的微管,用于装配子代病毒结构,并对细胞核周围进行维护。用MA104细胞培养轮状病毒时,在细胞周期中的S期分别发现NSP3、NSP5和VP2 3种轮状病毒蛋白,研究数据表明MA104细胞在细胞周期的S期和RV复制之间有很强的相关性。Bar-Magen等[18]阐明纯化的重组NSP5具有Mg2+依赖性和ATP特异性三磷酸酶活性,其产生游离ADP和Pi;ATPase活性与低水平的NSP5磷酸化相关,表明ATP水解与NSP5自身激活素活性之间可能存在联系。诱变显示细胞酪蛋白激酶样酶对NSP5高磷酸化所需的关键残基(Ser67)在NSP5的ATPase或自身激活活性中无作用。通过其NDP激酶活性,NSP2八聚体可以通过为NSP5的自激酶活性和与NSP5相关的细胞激酶产生恒定的ATP分子来支持NSP5磷酸化。
病毒感染细胞的免疫共沉淀实验得出,除NSP2与NSP5相互作用,VP1、VP2以及VP6与NSP5也存在相互作用。研究发现,NSP5与其他非结构蛋白无法接触到核心颗粒的dsRNA,与VP2、VP6的存在有关;而且发现轮状病毒感染细胞2至3 h后才能在细胞质内检测到病毒抗原。并在病毒基质支架中发现有4个结构蛋白VP1、 VP2、 VP3和VP6和非结构蛋白NSP2、 NSP5存在[11, 13, 19]。
NSP5的表达量是轮状病毒感染过程中判定的关键因素,并根据NSP5结构特征进行鉴别和分析[20];在病毒感染细胞的不同时期,通过NSP5结构的变化,可以作为轮状病毒在感染、复制和包装过程中的一种追踪蛋白,进而探讨轮状病毒感染后,细胞不同结构区域的变化进而调控轮状病毒的感染[21-24];接下来可以在病毒感染、复制、包装和释放的过程中,探讨研究各种蛋白间的相互作用方式,以便调控RV的感染。通过研究轮状病毒NSP5的作用,可以从致病机制中开发新的治疗方法,这将对新疫苗的开发有重要意义,也将会有助于控制和消除轮状病毒,从而控制和消除由轮状病毒引起的对人类健康的危害。
